Kamagra repose sur le sildénafil comme principe actif, avec un mode d’action identique à celui du Viagra. La forme galénique en gel oral permet une absorption plus rapide et une concentration plasmatique maximale plus précoce que les comprimés. Le mécanisme implique l’inhibition compétitive de la PDE5, entraînant une relaxation musculaire lisse locale et une vasodilatation ciblée. La demi-vie courte, environ 4 heures, limite la durée d’action. L’élimination se fait après métabolisme hépatique, impliquant majoritairement le CYP3A4. L’incidence d’effets indésirables comprend céphalées, rougeurs et congestion nasale, de façon transitoire. Dans les comparatifs pharmacologiques, acheter kamagra sans ordonnance est associé aux présentations galéniques alternatives disponibles.
The citric acid (krebs, tca) cycle
The Citric Acid (Krebs, TCA) Cycle Step 1: Condensation
In step 1 of the Krebs cycle, the two-carbon compound, acetyl-S-CoA, participates in a condensation reaction with the four-carbon compound, oxaloacetate, to produce citrate:
• This reacion is moderately exergonic. Thermodynamically, the equilibrium is in
favor of the products. Thus, this is considered to be the first committed step of the Krebs cycle
• Being the first committed step, this is a likely step to have some kind of
regulatory control mechanism (which will effectively regulate the entire cycle)
• The Krebs cycle is also known as the citric acid cycle. Citrate is a tricarboxylic
acid, and the Krebs cycle is also known as the tricarboxylic acid (or TCA) cycle
Step 2. Isomerization of Citrate
As we will see later on in the Krebs cycle, there will be a decarboxylation reaction.
• Such decarboxylation reactions usually involve α- (or β-) keto acids • The hydroxyl group of citrate can be oxidized to yield a keto group, but to form
an α-keto acid it needs to be adjacent to one of the terminal carboxyl groups
Thus, step 2 involves moving the hydroxyl group in the citrate molecule so that we can later form an α-keto acid
• This process involves a sequential dehydration and hydration reaction, to form the
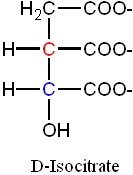
D-Isocitrate isomer (with the hydroxyl group now in the desired α- location), with cis-Aconitase as the intermediate
• A single enzyme, Aconitase, performs this two-step process:
• This reaction is endergonic, so the equilibrium is in favor of the reactants and not
the desired product. However, the exergonic character of the next reaction in the cycle helps shift the equilibrium of this reaction towards the right.
• There are two asymmetric centers in the D-Isocitrate molecule. Eeach can adopt
either the L- or D- rotamer, thus there are 4 possible isomers of this molecule
• Aconitase only produces the single form of Isocitrate (D-Isocitrate). Thus,
Aconitase is a stereospecific enzyme
Note: the stereospecificity of Aconitase was established by introducing carboxyl- labeled Acetate into the Krebs cycle. The conversion of Acetate into Acetyl-CoA can subsequently result in the labeling of Citrate. Although Citrate is a symmetric molecule, the labeled carboxyl-group always ends up on the γ- carbon group in D- Isocitrate Step 3: Generation of CO2 by an NAD+ linked enzyme
• The Krebs cycle contains two oxidative decarboxylation steps; this is the first one • The reaction is catalyzed by the enzyme Isocitrate dehydrogenase
• The reaction involves dehydrogenation to Oxalosuccinate, an unstable
intermediate which spontaneously decarboxylates to give α-Ketoglutarate
• The reaction is exergonic, with a ΔG0' = -20.9 kJ/mol. This helps drive the
preceding (endergonic) reaction in the cycle
• In addition to decarboxylation, this step produces a reduced nicotinamide adenine
dinucleotide (NADH) cofactor, or a reduced nicotinamide adenine dinucleotide phosphate (NADPH) cofactor
• If the NAD+ cofactor is reduced, then the D-Isocitrate must be oxidized when
forming α-Ketoglutarate. Thus, this step is referred to as an oxidative decarboxylation step Step 4: A Second Oxidative Decarboxylation Step
• This step is performed by a multi-enzyme complex, the α-Ketoglutarate
• The multi-step reaction performed by the α-Ketoglutarate Dehydration Complex
is analogous to the Pyruvate Dehydrogenase Complex, i.e. an α-keto acid undergoes oxidative decarboxylation with formation of an acyl-CoA
• Overall, this oxidative decarboxylation step is more exergonic than the first
Summary of Kreb cycle reactions up to this point Two carbons have been added to Oxaloacetate by the action of Citrate Synthase (and Acetyl-CoA)
• Two carbons have been lost as CO2 by oxidative decarboxylation steps • Two oxidized NAD+ cofactors have been reduced to NADH • Due to the stereospecific action of Aconitase, the two carbons added are not the
same two carbons lost in the oxidative decarboxylation steps
In the remaining steps of the Krebs cycle, the Succinyl-CoA is converted back into the original substrate for the cycle: OxaloacetateStep 5: Substrate-Level Phosphorylation
Succinyl-CoA is a high potential energy molecule. The energy stored in this molecule is used to form a high energy phosphate bond in a Guanine nucleotide diphosphate (GDP) molecule:
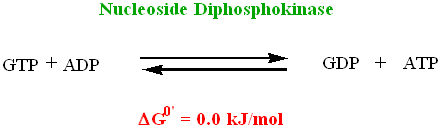
• Most of the GTP formed is used in the formation of ATP, by the action of
• In plants and bacteria ATP is formed directly in the Succinyl-CoA Synthase
catalyzed reaction by phosphorylation of ADP directly. In animals, GDP is the substrate in the reaction with formation of GTP (which is then used to form ATP by Nucleoside Diphosphokinase)
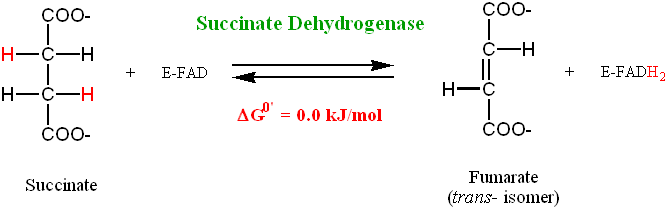
Step 6: Flavin-Dependent Dehydrogenation
The Succinate produced by Succinyl CoA-Synthetase in the prior reaction needs to be converted to Oxaloacetate to complete the Krebs cycle.
• Both Succinate and Oxaloacetate are 4-carbon compounds • The first step in the conversion is the dehydrogenation of Succinate to yield
• In this reaction a C-C bond is being oxidized to produce a C=C bond. This
oxidation is energetically more costly than oxidizing a C-O bond.
• The redox coenzyme for this reaction is therefore FAD, rather than NAD+ (FAD is
a more powerful oxidizing agent compared to NAD+)
• FAD is covalently bound to the Succinate Dehydrogenase molecule (via a
• The FADH2 has to be oxidized for the enzyme activity to be restored. This
oxidation occurs via interaction with the mitochondrial electron transport system (bound to mitochondrial inner membrane).
• Succinate Dehydrogenase is tightly bound to the mitochondrial inner membrane • Succinate Dehydrogenase is stereo-specific: the trans- isomer (Fumarate) is
produced and not the cis- isomer (Maleate)
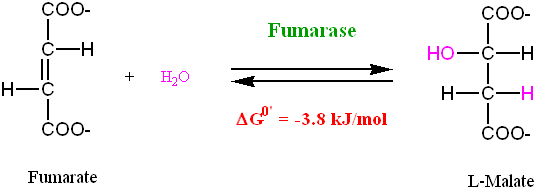
Step 7: Hydration of a Carbon-Carbon Double Bond
Fumarate undergoes a stereo-specific hydration of the C=C double bond, catalyzed by Fumarate Hydratase (also known as Fumarase), to produce L-Malate:
• Fumarase is a stereo-specific enzyme: it will only hydrate Fumarate, it will not
hydrate Maleate. Furthermore, the enzyme can not use D-Malate as a substrate in the reverse reaction
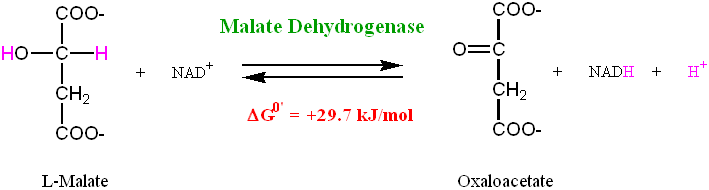
Step 8: A Dehydrogenation Reaction that will Regenerate Oxaloacetate
L-Malate (Malate) is dehydrogenated to produce Oxaloacetate by the enzyme Malate Dehydrogenase
• This is a highly endergonic reaction (ΔG0' = +29.7 kJ/mol) and so the equilibrium
strongly favors the reactants over the products.
• However, the next step in the Krebs cycle (i.e. the first step in the process) is the
highly exergonic reaction (ΔG0' = -32.2 kJ/mol) catalyzed by Citrate Synthase and this keeps the levels of Oxaloacetate low (<10-6 M), thus allowing the above reaction to proceed
• The formation of Oxaloacetate completes the Krebs cycle
Stoichiometry and Energetics of the Citric Acid Cycle Reaction (kJ/mol)
Acetyl-CoA + Oxaloacetate + H2O Ö Citrate + CoA-SH + H+
cis-Aconitase + H2O Ù Isocitrate
Isocitrate + NAD+ Ù α-Ketoglutarate + CO2 + NADH
α-Ketoglutarate + NAD+ + CoA-SH Ù Succinyl-CoA + CO2 + NADH α-Ketoglutarate
Succinyl-CoA + Pi + GDP Ù Succinate + GTP + CoA-SH
L-Malate + NAD+ Ù Oxaloacetate + NADH + H+
NET: -44.8
Acetyl-CoA + Oxaloacetate + H2O Ö Citrate + CoA-SH + H+
cis-Aconitase + H2O Ù Isocitrate
Isocitrate + NAD+ Ù α-Ketoglutarate + CO2 + NADH
α-Ketoglutarate + NAD+ + CoA-SH Ù Succinyl-CoA + CO2 + NADH
Succinyl-CoA + Pi + GDP Ù Succinate + GTP + CoA-SH
L-Malate + NAD+ Ù Oxaloacetate + NADH + H+
Acetyl-CoA + 2H2O + 3NAD+ + Pi + GDP + FAD Ö 2CO2 + 3NADH + GTP + CoA-SH
One turn of the citric acid cycle generates:
• One high-energy phosphate through substrate-level phosphorylation • Three NADH • One FADH2
Catabolism of Glucose through Glycolysis and the Krebs Cycle
• Each molecule of Glucose produces two molecules of Pyruvate
Glucose + 2NAD+ + 2ADP + 2Pi Ö 2Pyruvate + 2NADH + 2H+ + 2H2O +2ATP
• Action of Pyruvate Dehydrogenase on Pyruvate:
Pyruvate + CoA-SH + NAD+ Ö CO2 + Acetyl-CoA + NADH
• The overall catabolism of Glucose to 2 Pyruvate molecules:
Glucose + 2NAD+ + 2ADP + 2Pi Ö 2Pyruvate + 2NADH + 2H+ + 2H2O +2ATP
2Pyruvate + 2CoA-SH + 2NAD+ Ö 2CO2 + 2Acetyl-CoA + 2NADH
Glucose + 4NAD+ + 2ADP + 2CoA-SH + 2Pi Ö 2CO2 + 2Acetyl-CoA + 4NADH + 2H+
• The GTP formed in the animal Succinyl-CoA Synthetase reaction in the Krebs
cycle is readily converted to ATP (by Nucleoside Diphosphokinase)
2Acetyl-CoA + 4H2O + 6NAD+ + 2Pi + 2ADP + 2FAD Ö 4CO2 + 6NADH + 2ATP +
Glucose + 4NAD+ + 2ADP + 2CoA-SH + 2Pi Ö 2CO2 + 2Acetyl-CoA + 4NADH + 2H+
Glucose + 10NAD+ + 4ADP + 2H2O + 4Pi + 2FAD Ö 6CO2 + 10NADH + 4ATP +
Yield of ATP
At this point the yield of ATP is 4 moles per mole of Glucose as it passes through the Krebs cycle
• This is not much more than the 2 moles which would have been produced from
• However, NADH and FADH2 are energy rich molecules • Their oxidation is highly exergonic and is coupled with the production of ATP
• Oxidation of 1 mole NADH produces 3 moles ATP • Oxidation of 1 mole FADH2 produces 2 moles ATP • Thus total ATP yield = (10 x 3) + (2 x 2) + 4 = 38 moles ATP per mole Glucose
Prepared by Dr Hana M Gashlan
Die Gyn-Praxis Ihre kompetenten Frauenärzte Publikationen Lange, R. Plasmakinetik von ³H-Testosteron beim Hund: Der Effekt von Prolactin und Bromocriptin auf die Androgenelimination aus dem Plasma `ìn vivo´. Inauguraldissertation, Med.Fachbereiche, Universität Mainz 1977 Arenz,R., R.Lange : Bericht über eine primäre Abdominalgravidität, Arenz,R., R.Lange: Aufschlüss
A novel two-step kit for topical treatment of tinea pedis –an open studyA Shemer,† MH Grunwald,‡ B Davidovici,§ N Nathansohn,† B Amichai†,*†Department of Dermatology, Sheba Medical Center, Tel-Hashomer, ‡Soroka University Medical Center, Ben-Gurion University,Beer-Sheva, and §Dermatology Unit, Kaplan Medical Center, Rehovot, Israel*Correspondence: B Amichai. E-mail: boazam@clali
 The Citric Acid (Krebs, TCA) Cycle
The Citric Acid (Krebs, TCA) Cycle  Step 1: Condensation
Step 1: Condensation 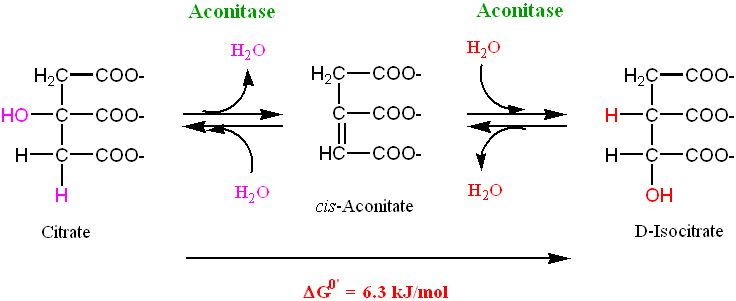
 • This reaction is endergonic, so the equilibrium is in favor of the reactants and not
the desired product. However, the exergonic character of the next reaction in the cycle helps shift the equilibrium of this reaction towards the right.
• There are two asymmetric centers in the D-Isocitrate molecule. Eeach can adopt
either the L- or D- rotamer, thus there are 4 possible isomers of this molecule
• Aconitase only produces the single form of Isocitrate (D-Isocitrate). Thus,
Aconitase is a stereospecific enzyme
Note: the stereospecificity of Aconitase was established by introducing carboxyl-
• This reaction is endergonic, so the equilibrium is in favor of the reactants and not
the desired product. However, the exergonic character of the next reaction in the cycle helps shift the equilibrium of this reaction towards the right.
• There are two asymmetric centers in the D-Isocitrate molecule. Eeach can adopt
either the L- or D- rotamer, thus there are 4 possible isomers of this molecule
• Aconitase only produces the single form of Isocitrate (D-Isocitrate). Thus,
Aconitase is a stereospecific enzyme
Note: the stereospecificity of Aconitase was established by introducing carboxyl-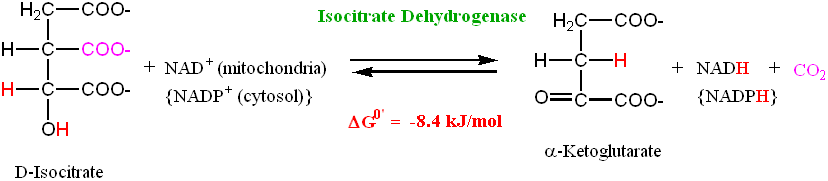
 • The reaction involves dehydrogenation to Oxalosuccinate, an unstable
intermediate which spontaneously decarboxylates to give α-Ketoglutarate
• The reaction is exergonic, with a ΔG0' = -20.9 kJ/mol. This helps drive the
preceding (endergonic) reaction in the cycle
• In addition to decarboxylation, this step produces a reduced nicotinamide adenine
dinucleotide (NADH) cofactor, or a reduced nicotinamide adenine dinucleotide phosphate (NADPH) cofactor
• If the NAD+ cofactor is reduced, then the D-Isocitrate must be oxidized when
forming α-Ketoglutarate. Thus, this step is referred to as an oxidative
• The reaction involves dehydrogenation to Oxalosuccinate, an unstable
intermediate which spontaneously decarboxylates to give α-Ketoglutarate
• The reaction is exergonic, with a ΔG0' = -20.9 kJ/mol. This helps drive the
preceding (endergonic) reaction in the cycle
• In addition to decarboxylation, this step produces a reduced nicotinamide adenine
dinucleotide (NADH) cofactor, or a reduced nicotinamide adenine dinucleotide phosphate (NADPH) cofactor
• If the NAD+ cofactor is reduced, then the D-Isocitrate must be oxidized when
forming α-Ketoglutarate. Thus, this step is referred to as an oxidative 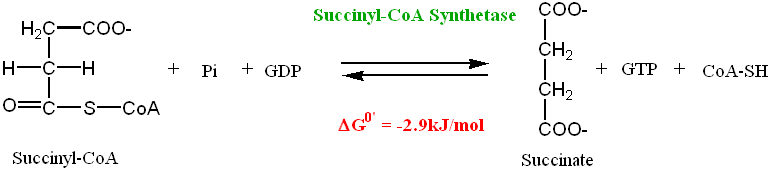
 • Fumarase is a stereo-specific enzyme: it will only hydrate Fumarate, it will not
hydrate Maleate. Furthermore, the enzyme can not use D-Malate as a substrate in the reverse reaction
• Fumarase is a stereo-specific enzyme: it will only hydrate Fumarate, it will not
hydrate Maleate. Furthermore, the enzyme can not use D-Malate as a substrate in the reverse reaction