Kamagra repose sur le sildénafil comme principe actif, avec un mode d’action identique à celui du Viagra. La forme galénique en gel oral permet une absorption plus rapide et une concentration plasmatique maximale plus précoce que les comprimés. Le mécanisme implique l’inhibition compétitive de la PDE5, entraînant une relaxation musculaire lisse locale et une vasodilatation ciblée. La demi-vie courte, environ 4 heures, limite la durée d’action. L’élimination se fait après métabolisme hépatique, impliquant majoritairement le CYP3A4. L’incidence d’effets indésirables comprend céphalées, rougeurs et congestion nasale, de façon transitoire. Dans les comparatifs pharmacologiques, acheter kamagra sans ordonnance est associé aux présentations galéniques alternatives disponibles.
Thoraxjnl-2011-200017 1.6
Thorax Online First, published on September 15, 2011 as 10.1136/thoraxjnl-2011-200017
Identification of FGF7 as a novel susceptibility locusfor chronic obstructive pulmonary disease
John M Brehm,1 Koichi Hagiwara,2 Yohannes Tesfaigzi,3 Shannon Bruse,3Thomas J Mariani,4 Soumyaroop Bhattacharya,4 Nadia Boutaoui,1 John P Ziniti,5Manuel E Soto-Quiros,6 Lydiana Avila,6 Michael H Cho,5,7,8 Blanca Himes,5Augusto A Litonjua,5,7,8,9 Francine Jacobson,10 Per Bakke,11 Amund Gulsvik,11Wayne H Anderson,12 David A Lomas,13 Erick Forno,14 Soma Datta,5Edwin K Silverman,5,7,8,15 Juan C Celedo
Rationale Traditional genome-wide association studies
(GWASs) of large cohorts of subjects with chronic
obstructive pulmonary disease (COPD) have successfully
identified novel candidate genes, but several other
< Can information from isolated populations
plausible loci do not meet strict criteria for genome-wide
improve our ability to detect novel genetic
significance after correction for multiple testing.
variants in genome-wide association studies
Objectives The authors hypothesise that by applying
unbiased weights derived from unique populations we
can identify additional COPD susceptibility loci.
Methods The authors performed a homozygosity
< We identified statistically significant polymor-
Children’s Hospital of Pittsburghof UPMC, 4401 Penn Avenue,
haplotype analysis on a group of subjects with and
phisms in a novel chronic obstructive pulmonary
without COPD to identify regions of conserved
disease (COPD) gene (FGF7), which we repli-
homozygosity haplotype (RCHHs). Weights were
constructed based on the frequency of these RCHHs in
case versus controls, and used to adjust the p values
from a large collaborative GWAS of COPD.
Results The authors identified 2318 RCHHs, of which
(homozygosity haplotype analysis) for identi-
576 were significantly (p<0.05) over-represented in
fying genomic regions that are inherited froma common ancestor, and use this information to
cases. After applying the weights constructed fromthese regions to a collaborative GWAS of COPD, the
authors identified two single nucleotide polymorphisms
genetic variants that are associated withincreased risk of disease.
(SNPs) in a novel gene (fibroblast growth factor-7(FGF7)) that gained genome-wide significance by thefalse discovery rate method. In a follow-up analysis, bothSNPs (rs12591300 and rs4480740) were significantly
significant results after correction for multiple
associated with COPD in an independent population
testing. Weighting the results of GWASs according to
prior information (eg, from linkage studies) may
respectively). In another independent population,
significantly improve the power to detect associations
increased lung tissue FGF7 expression was associated
that do not meet genome-wide (GW) significance.4
with worse measures of lung function.
Homozygosity mapping is a promising technique
Conclusion Weights constructed from a homozygosity
to identifying regions of the genome that are more
haplotype analysis of an isolated population successfully
identify novel genetic associations from a GWAS on
Although initially developed to identify rare
a separate population. This method can be used to
susceptibility mutations for monogenic traits in
identify promising candidate genes that fail to meet strict
families,5 homozygosity mapping has recently been
successfully applied to the study of complexdiseases.6 7 While techniques vary, the conceptunderlying all homozygosity haplotype (HH)methods is that regions of homozygosity are more
likely to contain disease-susceptibility loci in
affected subjects than in unaffected individuals.8
(GWASs) have identified novel susceptibility loci for
complex diseases such as chronic obstructive
morphism (SNP) arrays, Miyazawa et al developed
pulmonary disease (COPD).1e3 Because the effect
a novel variation of homozygosity mapping that
tests whether multiple subjects share the same
variants is modest, GWASs of complex diseases
require large sample sizes to achieve statistically
constructed a region of conserved homozygosity
Brehm JM, Hagiwara K, Tesfaigzi Y, et al. Thorax (2011). doi:10.1136/thoraxjnl-2011-200017
Copyright Article author (or their employer) 2011. Produced by BMJ Publishing Group Ltd (& BTS) under licence.
haplotype (RCHH) that reflects the transmission of the haplotype
multicentre ECLIPSE study.12 All controls were current or former
from a founder population. In theoretical simulations, this
smokers with normal spirometry, and all cases with COPD had
method was shown to be a viable method to detect disease-
moderate to very severe disease according to the Global Initiative
susceptibility loci in recently admixed populations.9 We hypoth-
for Chronic Obstructive Lung Disease classification.13
esised that application of this method to a genetic isolate in CostaRica would result in detection of an over-representation of regions
of conserved homozygosity in subjects affected with COPD
The top SNPs in novel genes were replicated in a cohort of 1845
compared with unaffected subjects. In this report, we first identify
smoking adults in New Mexico, 424 (23%) of whom were
regions of conserved homozygosity in Costa Ricans and then
classified with COPD based on an FEV1/FVC ratio below the
show that weights derived from these regions can be applied to
fifth percentile of the predicted value, also referred to as the
GWASs in non-isolated populations to identify novel disease-
lower limit of normal.14 Of the 1845 participants, 1411 (77%)
susceptibility loci for COPD. Using this approach, we identify
were Caucasian and 313 (17%) were Hispanic. The protocols for
a novel COPD candidate gene (fibroblast growth factor-7 (FGF7)).
subject recruitment and data collection for the Lovelace SmokersCohort have been previously described in detail.15 The two SNPs
(rs12591300 and rs4480740) were genotyped by allelic discrimi-
nation using Taqman assay (Applied Biosystems, Foster City,
The primary study population consisted of 58 subjects with
California, USA). The caseecontrol association analysis was first
COPD (cases) and 57 subjects without COPD (controls) in the
performed in all subjects, and then separately in Caucasians and
Genetic Epidemiology of COPD in Costa Rica study. Cases were
Hispanics. All analyses were adjusted for age, gender and pack-
recruited from patients attending four adult hospitals in San José
years of cigarette smoking; the analysis of all subjects was
(Costa Rica) and their affiliated clinics, and through newspaper
additionally adjusted for self-declared ethnicity.
advertisements. Control subjects were recruited from individualsattending a smoking-cessation clinic at the Institute for Phar-
maco-dependency in San José, and through newspaper adver-
For the top novel candidate genes, we examined the correlation of
tisements. To ensure their descent from the founder population
gene expression in lung tissue with COPD intermediate pheno-
of the Central Valley of Costa Rica (which is predominantly of
types (FEV1 and FEV1/FVC ratio) in a previously published
Spanish and Native American ancestry), all participants were
COPD biomarker discovery study.16 This cohort consists of 56
required to have at least six great-grandparents born in the
subjects with varying degrees of obstruction who underwent
Central Valley. Additional inclusion criteria for cases were ages
lung resection for a solitary pulmonary nodule. RNA expression
profiling was completed using the Affymetrix U133 Plus 2.0 array,
e71 years, physician-diagnosed COPD, $10 pack-years of
cigarette smoking, a forced expiratory volume in one second
as previously described.16 Expression correlation with quantita-
tive phenotypes was conducted as previously described.16
1) #65% predicted and an FEV1/forced vital capacity (FVC)
ratio of #70% after bronchodilator administration (180 mg ofalbuterol by metered dose inhaler). Controls were recruited on
the basis of the same criteria for age and smoking history, but
they had to have no physician-diagnosed COPD and normal
RCHHs were identified using the method described by Miyazawa
spirometry. Exclusion criteria for cases and controls included
et al.9 In brief, for any given individual all heterozygous SNPs
history of chronic pre-existing chronic lung disease (eg, bron-
were ignored and the SNP location was scored with the value of
chiectasis) and severe a-1-antitrypsin deficiency (for cases),
the allele for that subject. Subjects are compared only across SNPs
based on molecular phenotyping. The baseline characteristics of
that are scored. RCHHs are defined by runs of SNPs that share the
this cohort are in listed in the online supplementary table 2.
same allele at the homozygous locations across multiple subjects,
Written consent was obtained from participating subjects.
ignoring heterozygous SNPs. The size of the shared segments
The study was approved by the institutional review boards of
between any two individuals was set at 3.0 cM (roughly and
the Hospital Nacional de Niños (San José, Costa Rica), Partners
approximately three million base pairs), which in theoretical
Healthcare System (Boston, Massachusetts, USA), and partici-
work conducted by Miyazawa et al9 reduced the false positive
pating National Emphysema Treatment Trial (NETT), Evalua-
and false negative rates of discovery. A theoretical ancestral
tion of COPD Longitudinally to Identify Predictive Surrogate
segment was then constructed from the largest subgroup of
Endpoints (ECLIPSE) and Norway centres.
subjects sharing a particular RCHH (see online supplementaryfigure 1). While any two subjects must have at least 3.0 cM of
sharing, the size may be much smaller when comparing across
High-density SNP genotyping was performed using the Illumina
multiple subjects (online supplementary figure 2). If more than
Quad 610 platform at the Channing Laboratory, Boston,
one ancestral region is identified at a particular chromosomal
Massachusetts, USA. Cases and controls were randomly
location, the region shared by the most number of subjects is used
distributed among batches, and each batch contained a replicate
(online supplementary figure 3). The total number of cases and
sample. All subjects had an SNP call rate >95%. After quality
controls sharing this ancestral allele is used to calculate a p value
control measures (see online supplementary table 1), a total of
based on a standard normal distribution.
558 929 SNPs were acceptable for analysis.
For the primary analysis of the collaborative COPD cohort,
logistic regression analysis was performed under an additive
Collaborative COPD cohorts for the primary GWAS
genetic model for each SNP, adjusting for age, pack-years of
Three populations with a total of 2940 cases and 1380 controls
smoking and the first 16 principal components (to adjust for
were used for the primary GWAS: (1) subjects in a caseecontrol
population stratification). The p values from all RCHHs iden-
study of COPD in Norway (838 cases and 791 controls)3; (2)
tified in Costa Rica were then used to construct a cumulative
subjects in the NETT (366 cases) and the Normative Aging Study
weight for each SNP from the recent GWAS of COPD in the
(414 controls)10 11 and (3) 1736 cases and 175 controls from the
combined cohort of Norway, ECLIPSE and NETTeNormative
Brehm JM, Hagiwara K, Tesfaigzi Y, et al. Thorax (2011). doi:10.1136/thoraxjnl-2011-200017
Aging Study using the method developed by Roeder et al.4
there was an RCHH containing FAM13A identified in the Costa
Briefly, the weighting method utilises prior information (in this
Rican cohort, it was only seen in one case and no controls.
case, the p value representing the degree of over-representationof a region of the genome in cases versus controls) to upweight
or downweight p values from an association study (in this case,
The top two SNPs in or near FGF7 were genotyped in the 1845
the GWAS of COPD in the collaborative cohort). In order to
smoking adults in the Lovelace Smokers Cohort. The minor
maintain an overall a level of 0.05, the assigned weights across
alleles of both SNPs conferred increased odds for COPD in the
the genome average to 1. For this study, SNPs that did not fall
whole population in the same direction as the original collabo-
inside of an RCHH (and therefore did not have a p value) were
rative COPD cohort (table 2). Among the Hispanic subgroup,
assigned a p value of 1 (and therefore a weight approaching
the effect size was larger and in the same direction for both
zero). This is a more conservative approach than excluding these
SNPs, but only rs12591300 showed a significant association with
SNPs from consideration. The method then calculates a false-
discovery rate (FDR) using the method described by Benjaminiand Hochberg17 to correct for multiple testing.
The RCHHs were created and compared with HHAnalysis
Our previous studies indicate that gene expression patterns
(available at http://www.hhanalysis.com). Association analysis
associated with quantitative, intermediate COPD phenotypes
was performed using PLINK V.1.07 (http://pngu.mgh.harvard.
are most informative for the discovery of disease-associated
edu/purcell/plink). The weighting procedure was performed
genes.16 18 19 We examined disease-associated expression
using software developed by Roeder et al4 (http://wpicr.wpic.
patterns for our novel candidate genes in a previously published
pitt.edu/wpiccompgen/). All other statistical analysis was
GW expression data set from 56 subjects with varying degrees
performed using R V.2.9.0 (http://www.R-project.org).
of airflow obstruction (assessed by spirometric measures of lungfunction (FEV1 and FEV1/FVC ratio)).16 Expression of FGF7 (asdefined by multiple and independent probe sets) was signifi-
cantly negatively correlated with both FEV
Identification of RCHHs in Costa Rica and construction of
1/FVC ratio (nominal p value <0.01), indicating
increased expression associated with increased disease severity.
In total, 2318 RCHHs were identified in the Costa Rican cohort.
Expression in COPD subjects was increased compared with
Of these 2318 regions, 576 were significantly (p<0.05) over-
control subjects, but the difference was not statistically
represented in cases compared with controls; none of the regions
significant. PSMA4 expression was not correlated with lung
were significantly more frequent in controls than cases. The
function and was not differentially expressed in cases versus
median size of the significant regions was 105 kb, and the largest
was 7.2 Mb. Online supplementary table 3 shows the top 20 pvalues representing 100 RCHHs in Costa Rica.
Each SNP in the combined collaborative COPD cohort was
then mapped to an RCHH and assigned the p value of the whole
While successful in identifying novel candidate genes, GWASs of
region. SNPs that did not map to an RCHH were assigned a p
complex traits are unlikely to identify all potential common
value of 1. The mapped p values across all genotyped SNPs were
disease-susceptibility variants because of limited power if strict
then used to create weights using a cumulative distribution
criteria for GW significance are applied. In the absence of a very
function. The algorithm is constructed so that the mean weight
large sample size, novel methods are needed to identify disease-
across all SNPs is 1: some SNPs are upweighted and a much
susceptibility variants not meeting GW significance. We identi-
larger fraction is downweighted. The nominal p value is divided
fied RCHHs for COPD in a GW caseecontrol study in Costa
by the weight to obtain the weighted p value.
Rica. After applying a weighting method based on the degree ofsignificance of these regions to a GWAS of COPD cases and
controls of European descent, we identified two SNPs in a novel
We applied the weights derived from the HH analysis above to
candidate gene for COPD (FGF7) and demonstrated that several
reanalyse GW genotypic data in a cohort of subjects of European
SNPs in the previously identified candidate genes IREB2 and
descent that was previously employed for a traditional GWAS of
CHRNA3 met GW criteria for statistical significance. An SNP in
COPD. After weighting, 14 SNPs were significant at an FDR-
another novel gene (PSMA4) was GW significant after
corrected a of 0.05. The top five SNPs from the unweighted
weighting. However, expression of PSMA4 in the lung was not
GWAS retained their original ranks, but several SNPs that did
associated with COPD phenotypes, and thus the observed
not achieve GW significance in the traditional GW association
association is likely due to linkage disequilibrium with the
analysis became more statistically significant and moved higher
nearby genes CHRNA3 and IREB2. We then replicated the two
in the list (table 1). Of these SNPs, those in the gene for FAM13A
FGF7 SNPs in an independent cohort of smoking adults, and
were identified in the original analysis of the GWAS,1 and SNPs
showed that they are both significantly associated in the same
in IREB218 and CHRNA33 have been implicated in COPD
direction with COPD. Notably, the effect sizes in Hispanics are
affection status in prior candidate-gene and GWASs. Two of the
larger than in the overall cohort, suggesting that these alleles
other SNPs lie in two novel candidate genes for COPD, FGF7
confer greater risk in this population. This Hispanic population
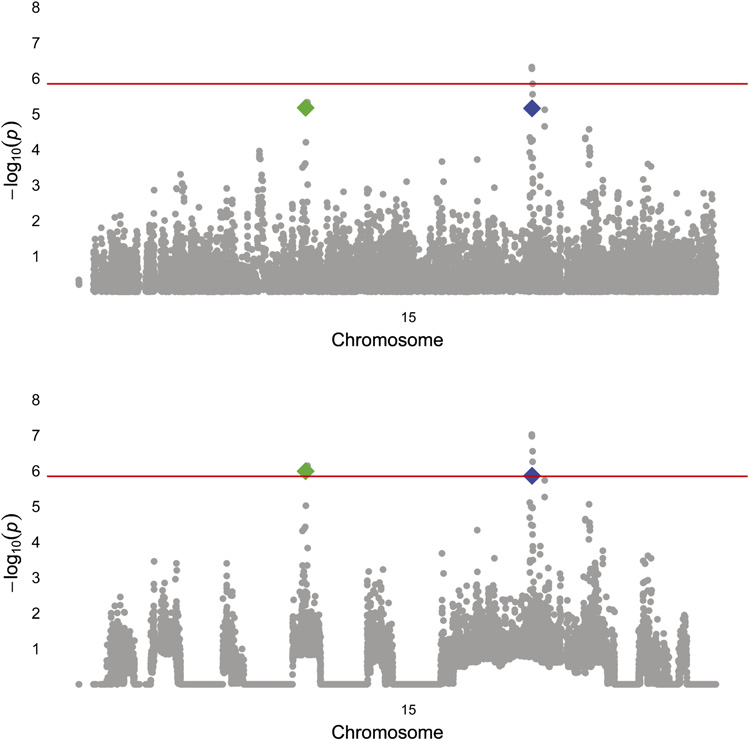
and proteasome subunit, a-type, 4 (PSMA4) (figure 1). The
in New Mexico has a similar proportion of European and Native
RCHH in Costa Rica that contains FGF7 was present in seven
American ancestry as the Costa Rican cohort,20 21 so another
cases and no controls, and the RCHH containing PSMA4 was
likely possibility is that patterns of linkage disequilibrium may
present in five cases and no controls.
be different between Hispanics and Caucasians in this genomic
The regions containing the genes CHRNA3 and IREB2 were
region, and that these SNPs are tagging a haplotype or func-
also over-represented in cases compared with controls (p<0.05),
tional SNP in the Hispanic subjects. Additionally, there was
and after weighting they were GW significant by FDR. While
a trend towards increased lung tissue expression of FGF7 in an
Brehm JM, Hagiwara K, Tesfaigzi Y, et al. Thorax (2011). doi:10.1136/thoraxjnl-2011-200017
FDR significant* results from weighted GWAS
*An FDR-corrected p value of 1.43Ee6 was used as the cut-off for genome-wide significance.
yResults previously published by Cho et al.1
zThe weighted p value is the original p value divided by the weight constructed from the RCHH (not shown).
xp Values for individual cohorts are the original, unweighted p values. COPD, chronic obstructive pulmonary disease; ECLIPSE, Evaluation of COPD Longitudinally to Identify Predictive Surrogate Endpoints; FDR, false-discovery rate; FGF7, fibroblast growth factor-7;GWAS, genome-wide association study; NAS, Normative Aging Study; NETT, National Emphysema Treatment Trial; PSMA4, proteasome subunit, a-type, 4; RCHH, region of conservedhomozygosity haplotype; SNP, single nucleotide polymorphism.
independent cohort of COPD subjects, in whom there was
There are several plausible methods for weighting chromo-
a significant negative correlation between FGF7 expression and
somal regions in GWAS, including upweighting previously
identified candidate genes, coding variants, exons and promoter
chromosome 15, before (top) and afterweighting. rs4480740 (Green) is in thegene FGF7 and rs2036534 (blue) is inthe promoter of PSMA4. The red lineindicates the FDR corrected a level forgenome-wide significance. FDR, false-discovery rate; FGF7, fibroblast growthfactor-7; PSMA4, proteasome subunit,
Brehm JM, Hagiwara K, Tesfaigzi Y, et al. Thorax (2011). doi:10.1136/thoraxjnl-2011-200017
Combined p values for replication of FGF7 SNPs
*Fisher’s combined p value using original two-sided p values. COPD, chronic obstructive pulmonary disease; FGF7, fibroblast growth factor-7; SNP, single nucleotide polymorphism.
regions. However, these weighting strategies work counter to
power of the algorithm to detect them. Genetic and historical
one of the strengths of a GWAS: its hypothesis-free nature.
data for the population of the Central Valley of Costa Rica
Using HHs as a weighting method avoids the pitfall of these
suggest that the first three assumptions are met. As in most
other weighting strategies because they are constructed using
association studies of complex disease, the effect size of a risk
a hypothesis-free method, so the weights are unbiased with
allele is likely small to moderate at most, and we expect that this
One of the main strengths of our study is that it shows the
Whereas other homozygosity mapping methods are primarily
power of using HH analysis in an isolated population to inves-
designed to detect recessive alleles, the HHAnalysis method
tigate common diseases. While our sample size was small,
instead uses homozygosity to identify ancestral regions inherited
Miyazawa et al9 have previously shown in simulated data that
from a common ancestor. These regions from a common ancestor
HH analysis has the ability to identify the region containing an
can harbour risk alleles that operate under recessive, dominant or
SNP inherited identity-by-descent from a distant common
additive models. However, the HHAnalysis algorithm would also
ancestor using only 45 cases and 45 controls. In our own
detect copy number variation that results in the deletion of
data, we were able to show that the previously identified
a single allele. While this may explain a fraction of the regions
candidate genes IREB2 and CHRNA3 fall within an RCHH that
identified, the top novel SNPs identified in FGF7 do not fall
is significantly over-represented in subjects with COPD. When
within known regions of copy number variation according to the
combined with results from a weighted GWAS in an indepen-
dent cohort with adequate sample size, we were able to show
In summary, we have shown that weights obtained from HH
that variants in these genes are significant after correction for
analysis in an isolated population can improve the power to
detect novel variants in GWAS in non-isolates. In addition to
Two novel genes are contained within significant regions of
confirming results for previously identified variants in IREB2
conserved homozygosity, and after weighting they are signifi-
and CHRNA3, we have identified variants in a novel candidate
cant by FDR correction. The first, FGF7, was identified in
gene (FGF7) for COPD. The validity of this gene is supported
cultured human embryonic lung fibrobasts,22 and plays a role in
by replication in an independent cohort of smoking adults, and
promoting wound healing23 and protecting airway epithelium
expression data showing consistent and significant patterns
from oxidant injury in mice.24 One of the SNPs identified in this
associated with COPD intermediate lung function phenotypes.
study (rs4480740) is in an intron of FGF7, and the other
Further analysis of these genes in the Costa Rican cohort
(rs12591300) lies immediately upstream of FGF7 in an intron of
and functional studies should yield insights into the causative
hypothetical protein LOC196951. In a GWAS of FEV1 in the
SNPs or haplotypes that underlie the associations identified in
British 1958 Birth Cohort, five out of the nine SNPs genotyped
in FGF7 were significantly (p<0.05) associated with differencesin lung function, although not the two SNPs identified in this
study.25 FGF7 has been shown to protect against oxidative stress
Division of Pediatric Pulmonary Medicine, Allergy, and Immunology, Children’s
Hospital of Pittsburgh of UPMC, Pittsburgh, Pennsylvania, USA
response specifically in the lung epithelium,24 so increases in
2Saitama Medical University Hospital and Institute, Saitama, Japan
expression associated with disease progression may indicate
3Lovelace Respiratory Research Institute, Albuquerque, New Mexico, USA
a greater burden of injury. A limitation of our study is the lack of
4University of Rochester Medical Center, Rochester, New York, USA5
experimental evidence for an effect(s) of the SNPs identified in
Channing Laboratory, Brigham and Women’s Hospital, Boston, Massachusetts, USA
Division of Pediatric Pulmonology, Hospital Nacional de Nin
on gene expression. We hypothesise that these SNPs cause
7Division of Pulmonary/Critical Care Medicine, Brigham and Women’s Hospital, Boston,
decreased expression of FGF7, which could affect antioxidant
mechanisms protecting against detrimental effects of cigarette
8Department of Medicine, Brigham and Women’s Hospital, Boston, Massachusetts,
smoking on the lung. Alternatively, FGF7 may play a role in
disease susceptibility through its role in epithelial development
Department of Medicine, Harvard Medical School, Boston, Massachusetts, USA
10Department of Radiology, Brigham and Women’s Hospital, Boston, Massachusetts,
during embryogenesis by influencing epithelial responses to
cigarette smoke. Since it is unclear whether increased FGF7
11Haukeland University Hospital and Institute of Medicine, University of Bergen,
expression is a marker of exposure to oxidant injury or a cause of
epithelial damage, further work must be done to characterise the
GlaxoSmithKline Research and Development, Research Triangle Park, North Carolina,
role of these SNPs on FGF7 expression.
13Cambridge Institute for Medical Research, University of Cambridge, Cambridge, UK
The HHAnalysis algorithm works best under certain
14Division of Pediatric Pulmonology, Department of Pediatrics, University of Miami,
assumptions, namely that (1) the risk alleles were introduced
into the population from a population of common ancestors
Center for Genomic Medicine, Brigham and Women’s Hospital, Boston,
within the last several hundred years, (2) the target population is
genetically isolated, (3) the number of common ancestors
Funding The Genetic Epidemiology of COPD in Costa Rica is supported by grant
introducing the risk allele is small and that (4) the risk of the
R01HL073373 from the National Heart, Lung, and Blood Institute. The NationalEmphysema Treatment Trial (NETT) is supported by contracts with the National Heart,
disease allele is moderate to high. Violations of these assump-
Lung, and Blood Institute (N01HR76101, N01HR76102, N01HR76103, N01HR76104,
tions reduce the theoretical expected size of the RCHH and/or
N01HR76105, N01HR76106, N01HR76107, N01HR76108, N01HR76109,
the association of the RCHH with disease, which reduces the
N01HR76110, N01HR76111, N01HR76112, N01HR76113, N01HR76114,
Brehm JM, Hagiwara K, Tesfaigzi Y, et al. Thorax (2011). doi:10.1136/thoraxjnl-2011-200017
N01HR76115, N01HR76116, N01HR76118, N01HR76119), the Centers for Medicare
Bell B, Rose CL, Damon A. The Normative Aging Study: an interdisciplinary and
and Medicaid Services (CMS) and the Agency for Healthcare Research and Quality
longitudinal study of health and aging. Aging Hum Dev 1972;3:5e17.
(AHRQ). The Norway cohort and the ECLIPSE study (http://clinicaltrials.gov identifier
Fishman A, Martinez F, Naunheim K, et al. A randomized trial comparing lung-
NCT00292552; GSK Code SCO104960) are funded by GlaxoSmithKline. The Lovelace
volume-reduction surgery with medical therapy for severe emphysema. N Engl J
Smokers Cohort is supported by funding from the State of New Mexico (appropriation
from the Tobacco Settlement Fund) and by grant RO1 ES015482 from the National
Vestbo J, Anderson W, Coxson HO, et al. Evaluation of COPD Longitudinally to
Institute of Environmental Health Sciences. We acknowledge use of genotype data
Identify Predictive Surrogate End-points (ECLIPSE). Eur Respir J 2008;31:869
from the British 1958 Birth Cohort DNA collection, funded by the Medical Research
Pauwels RA, Buist AS, Calverley PM, et al. Global strategy for the diagnosis,
Council grant G0000934 and the Wellcome Trust grant 068545/Z/02.
management, and prevention of chronic obstructive pulmonary disease. NHLBI/WHO
Global Initiative for Chronic Obstructive Lung Disease (GOLD) Workshop summary. Am J Respir Crit Care Med 2001;163:1256e76.
Ethics approval Institutional Review Board of University of Pittsburgh, Partners
Pellegrino R, Viegi G, Brusasco V, et al. Interpretative strategies for lung function
Health Care (Boston), participating NETT centres, Boston VA, Norway, Costa Rica, and
Sood A, Stidley CA, Picchi MA, et al. Difference in airflow obstruction betweenHispanic and non-Hispanic White female smokers. COPD 2008;5:274
Contributors Manuscript preparation: JMB, JCC; data analysis and study design:
Bhattacharya S, Srisuma S, Demeo DL, et al. Molecular biomarkers for
JMB, YT, SB, TJM, SB, NB, JPZ, MES-O, LA, MHC, BH, AAL, FJ, EF, SD, EKS, JCC;
quantitative and discrete COPD phenotypes. Am J Respir Cell Mol Biol
data collection: YT, TJM, MES, LA, AAL, PB, AG, WHA, DAL, EKS, JCC; statistical
Benjamini Y, Hochberg Y. Controlling the false discovery rate: a practical and
Provenance and peer review Not commissioned; externally peer reviewed.
powerful approach to multiple testing. J Roy Stat Soc B Stat Meth1995;57:289e300.
DeMeo DL, Mariani T, Bhattacharya S, et al. Integration of genomic and genetic
approaches implicates IREB2 as a COPD susceptibility gene. Am J Hum Genet
Cho MH, Boutaoui N, Klanderman BJ, et al. Variants in FAM13A are associated with
chronic obstructive pulmonary disease. Nat Genet 2010;42:200e2.
Demeo DL, Mariani TJ, Lange C, et al. The SERPINE2 gene is associated
Hirschhorn JN. Genomewide association studiesdilluminating biologic pathways.
with chronic obstructive pulmonary disease. Am J Hum Genet
Pillai SG, Ge D, Zhu G, et al. A genome-wide association study in chronic
Klimentidis YC, Miller GF, Shriver MD. Genetic admixture, self-reported ethnicity,
obstructive pulmonary disease (COPD): identification of two major susceptibility loci.
self-estimated admixture, and skin pigmentation among Hispanics and Native
Americans. Am J Phys Anthropol 2009;138:375e83.
Roeder K, Bacanu SA, Wasserman L, et al. Using linkage genome scans to
Morera B, Barrantes R, Marin-Rojas R. Gene admixture in the Costa Rican
improve power of association in genome scans. Am J Hum Genet 2006;78:243e52.
population. Ann Hum Genet 2003;67:71e80.
Lander ES, Botstein D. Homozygosity mapping: a way to map human recessive
Rubin JS, Osada H, Finch PW, et al. Purification and characterization of a newly
traits with the DNA of inbred children. Science 1987;236:1567e70.
identified growth factor specific for epithelial cells. Proc Natl Acad Sci U S A
Lencz T, Lambert C, DeRosse P, et al. Runs of homozygosity reveal highly penetrant
recessive loci in schizophrenia. Proc Natl Acad Sci U S A 2007;104:19942e7.
Werner S, Smola H, Liao X, et al. The function of KGF in morphogenesis of
Morrow EM, Yoo SY, Flavell SW, et al. Identifying autism loci and genes by tracing
epithelium and reepithelialization of wounds. Science 1994;266:819e22.
recent shared ancestry. Science 2008;321:218e23.
Ray P, Devaux Y, Stolz DB, et al. Inducible expression of keratinocyte growth factor
Reich DE, Schaffner SF, Daly MJ, et al. Human genome sequence variation and the
(KGF) in mice inhibits lung epithelial cell death induced by hyperoxia. Proc Natl Acad
influence of gene history, mutation and recombination. Nat Genet 2002;32:135e42.
Miyazawa H, Kato M, Awata T, et al. Homozygosity haplotype allows
http://www.b58cgene.sgul.ac.uk/ (accessed 8 Jan 2010).
a genomewide search for the autosomal segments shared among patients. Am J
Iafrate AJ, Feuk L, Rivera MN, et al. Detection of large-scale variation in the human
Brehm JM, Hagiwara K, Tesfaigzi Y, et al. Thorax (2011). doi:10.1136/thoraxjnl-2011-200017
Identification of FGF7 as a novel susceptibility locus for chronic obstructive pulmonary disease
John M Brehm, Koichi Hagiwara, Yohannes Tesfaigzi, et al. Thoraxdoi: 10.1136/thoraxjnl-2011-200017
Updated information and services can be found at:
Data Supplement
Published online September 15, 2011 in advance of the print journal. Email alerting
Receive free email alerts when new articles cite this article. Sign up in
the box at the top right corner of the online article.
Advance online articles have been peer reviewed, accepted for publication, edited andtypeset, but have not not yet appeared in the paper journal. Advance online articles are citable and establish publication priority; they are indexed by PubMed from initialpublication. Citations to Advance online articles must include the digital object identifier (DOIs) and date of initial publication.
Futter und Futtermanagement Richtiges Futter in der richtigen Menge zur richtigen Zeit ist die halbe Diabeteseinstellung. Das beste Insulin bewirkt keine guten Zuckerwerte, wenn falsch gefüttert wird. Zum Futter : Bei diabetischen Katzen füttert man proteinreich, moderat Fett, wenig Kohlenhydrate. Diätfutter sind nicht zwingend notwendig. Trockenfutter sollte man vermeiden, es enth�
MG42710K 1Q12 EMORY MEMBER GUIDE_v3_MG42710K 1Q12 EMORY M Medication Pocket Guide for Emory Members Please bring this guide with you on your next doctor visit. Medco manages your prescription drug benefit for your health plan. MG42710K 1Q12 EMORY MEMBER GUIDE_v3_MG42710K 1Q12 EMORY MMG42710K 1Q12 EMORY MEMBER GUIDE_v3_MG42710K 1Q12 EMORY M UNDERSTANDING YOUR PLAN The Emory Plan uses a d
 FDR significant* results from weighted GWAS
*An FDR-corrected p value of 1.43Ee6 was used as the cut-off for genome-wide significance.
FDR significant* results from weighted GWAS
*An FDR-corrected p value of 1.43Ee6 was used as the cut-off for genome-wide significance. Identification of FGF7 as a novel
Identification of FGF7 as a novel