Kamagra repose sur le sildénafil comme principe actif, avec un mode d’action identique à celui du Viagra. La forme galénique en gel oral permet une absorption plus rapide et une concentration plasmatique maximale plus précoce que les comprimés. Le mécanisme implique l’inhibition compétitive de la PDE5, entraînant une relaxation musculaire lisse locale et une vasodilatation ciblée. La demi-vie courte, environ 4 heures, limite la durée d’action. L’élimination se fait après métabolisme hépatique, impliquant majoritairement le CYP3A4. L’incidence d’effets indésirables comprend céphalées, rougeurs et congestion nasale, de façon transitoire. Dans les comparatifs pharmacologiques, acheter kamagra sans ordonnance est associé aux présentations galéniques alternatives disponibles.
Research.yerkes.emory.edu
Neuropsychopharmacology (2005), 1–9& 2005 Nature Publishing Group
Olanzapine-Induced Suppression of CocaineSelf-Administration in Rhesus Monkeys
Leonard L Howell*,1,2, Kristin M Wilcox1, Kimberly P Lindsey1 and Heather L Kimmel1
Yerkes National Primate Research Center, Emory University, Atlanta, GA, USA; 2Department of Psychiatry and Behavioral Sciences,
The neuropharmacological profile of the atypical antipsychotic, olanzapine, is consistent with a potentially useful medication for cocaine
abuse. The present study utilized an i.v. drug self-administration paradigm in nonhuman primates to obtain definitive evidence regarding
the effectiveness of olanzapine to modulate the reinforcing effects of cocaine. The effects of olanzapine were compared directly to those
of the neuroleptic, haloperidol. Rhesus monkeys (n ¼ 7) were trained to self-administer cocaine (0.03–0.3 mg/kg/injection) under a
second-order, fixed-interval 600-s schedule with fixed ratio 20 components. Experimental sessions comprised five consecutive fixed
intervals, each followed by a 1-min timeout. In drug-interaction experiments, a single dose of olanzapine (0.03–0.3 mg/kg) or haloperidol
(0.01–0.03 mg/kg) was administered i.v. 15 min presession for at least three consecutive sessions. In drug-substitution experiments,
different doses of olanzapine (0.01–0.1 mg/kg/injection) were substituted for cocaine until responding stabilized. Olanzapine caused dose-
related decreases in cocaine self-administration at pretreatment doses that had no overt behavioral effects indicative of sedation. A dose
of 0.1 mg/kg eliminated cocaine self-administration in all subjects. In contrast, doses of haloperidol that suppressed cocaine self-
administration induced marked sedation and catalepsy. Olanzapine failed to maintain self-administration behavior above saline extinction
levels over a range of unit doses. In vivo microdialysis experiments in a second group of awake rhesus monkeys (n ¼ 3) confirmed
previous reports in rodents that olanzapine effectively increases extracellular dopamine in ventral striatum. The dose of olanzapine that
markedly suppressed cocaine self-administration behavior increased dopamine to approximately 190% of control values. Lastly,
pretreatment with fluoxetine had no systematic effect on olanzapine-induced increases in striatal dopamine. The results indicate that
olanzapine can effectively suppress cocaine self-administration behavior in nonhuman primates at doses that enhance dopamine release
but do not maintain drug self-administration.
Neuropsychopharmacology advance online publication, 27 July 2005; doi:10.1038/sj.npp.1300828
Keywords: olanzapine; cocaine; dopamine; drug self-administration; microdialysis; nonhuman primates
like drugs for DAT correlate well with their potencies forsupporting self-administration behavior (Ritz et al, 1987;
Cocaine abuse remains a significant health concern, yet no
Bergman et al, 1989, Wilcox et al, 1999). In humans, a
effective pharmacotherapy is currently in clinical use.
significant correlation has been observed between DAT
Cocaine blocks the reuptake of monoamines including
occupancy and the intensity of subjective effects produced
dopamine, norepinephrine, and serotonin with approxi-
by intravenous cocaine (Volkow et al, 1997). Conversely,
mately equal potency (Heikkila and Manzino, 1984; Reith
dopamine antagonists can attenuate specific behavioral
et al, 1986; Kuhar et al, 1991). However, the acute
effects of cocaine in a surmountable manner including its
behavioral effects of cocaine in rodents and primates have
reinforcing effects (De Wit and Wise, 1977; Woolverton,
been linked most closely to increases in dopamine
1986; Hemby et al, 1996). Cocaine affects neurotransmission
associated with dopamine transporter (DAT) inhibition
in various brain dopamine systems leading to a variety of
(Wise, 1984; Ritz et al, 1987; Kleven and Woolverton, 1993;
behavioral effects, but the mesolimbic/mesocortical dopa-
Howell and Wilcox, 2001). The affinities of several cocaine-
mine system appears to be a critical mediator of thereinforcing effects of cocaine (Kuhar et al, 1991; Wise,
*Correspondence: Dr LL Howell, Yerkes National Primate Research
1998). Collectively, the results obtained in behavioral
Center, Emory University, 954 Gatewood Road NE, Atlanta, GA
studies provide compelling evidence that dopamine plays
30329, USA, Tel: þ 1 404 727 7730, Fax: þ 1 404 727 1266,
a major role in the neuropharmacology and addictive
E-mail: [email protected] 12 January 2005; revised 9 May 2005; accepted 7 June 2005
Online publication: 20 June 2005 at http://www.acnp.org/citations/
Given the obvious importance of dopaminergic mechan-
isms in the addictive properties of cocaine, the development
Olanzapine and cocaine self-administration
and use of compounds that target dopaminergic systems
Animals’ and were approved by the Institutional Animal
represents a reasonable approach for the pharmacological
Care and Use Committee of Emory University.
treatment of cocaine abuse. The neuropharmacologicalprofile of the atypical antipsychotic, olanzapine, is con-
sistent with a potentially useful medication. Olanzapine isan antagonist with high affinity for dopamine D
Each subject was prepared with a chronic indwelling venous
catheter under sterile surgical conditions using a technique
4-receptors (Moore et al, 1993; Bymaster et al, 1996, 1997),
and it appears to have mesolimbic selectivity (Moore et al,
described previously (Wilcox et al, 2002). Preoperative
1993) consistent with other atypical antipsychotics (Hand
antibiotics (Rocephin, 25 mg/kg or Cefazolin, 25 mg/kg)
et al, 1987). Interestingly, olanzapine induces significant
were given on the day of surgery to help prevent infection. A
elevations in extracellular dopamine in vivo in the
silicone catheter (0.65 mm ID, 1.75 mm OD; Access Techno-
prefrontal cortex, striatum, and nucleus accumbens of rats
logies, Skokie, IL) was implanted under a combination of
(Li et al, 1998; Zhang et al, 2000; Koch et al, 2004). In
Telazol (4.0 mg/kg) and isoflurane anesthesia using aseptic
behavioral studies, pretreatment with olanzapine produced
techniques. The proximal end of the catheter terminated in
dose-dependent decreases in cocaine self-administration
the vena cava above the right atrium, and the distal end was
in rats trained under a simple fixed-ratio schedule of
routed under the skin and attached to a subcutaneous
i.v. drug delivery (Meil and Schechter, 1997). Clozapine,
vascular access port (Access Technologies, Skokie, IL)
another atypical antipsychotic with a pharmacological
located in the center of the lower back. After surgery, the
subject was returned to its home cage and received
Bymaster et al, 1996), significantly attenuated the subjective
Banamine (1.0 mg/kg) every 6 h for 24 h postoperatively to
effects of cocaine in human cocaine abusers (Farren et al,
reduce pain and discomfort associated with surgery.
2000). Collectively, these studies suggest a potential thera-
Catheters were flushed daily with 100 U/ml heparinized
peutic role for olanzapine in the treatment of cocaine
saline to maintain patency. In experiments involving in vivo
abuse. Moreover, its antipsychotic actions could serve to
microdialysis, guide cannulae were implanted bilaterally
alleviate stimulant-induced psychotic-like symptoms that
into the caudate nucleus under sterile conditions. The
are not targeted by other medications currently under
positioning of the guide cannulae allowed for targeting of
the ventral striatum corresponding to the nucleus accum-
The present study determined the effectiveness of
bens. Preoperative antibiotics (Rocephin, 25 mg/kg) were
olanzapine in suppressing cocaine self-administration
given on the day of surgery to help prevent infection. The
behavior maintained by a complex schedule of i.v. drug
animals were sedated with Telazol (4.0 mg/kg) and main-
delivery in rhesus monkeys. The effects of olanzapine were
tained on isoflurane anesthesia during the surgery. The
compared directly to those of the neuroleptic, haloperidol,
subjects were positioned in a stereotaxic frame, and
in order to access the potential contribution of motor side
coordinates derived from MRI were used for accurate
effects. In addition, olanzapine was substituted for cocaine
probe placement. A trephine drill was used to make two
in drug self-administration experiments in order to
small burr holes directly above the ventral striatum, and the
characterize its reinforcing properties in monkeys with a
guide cannulae were inserted to the appropriate depth.
history of cocaine use. The drug-substitution procedure is a
Teflon screws attached to the skull were used to anchor
well-validated predictor of abuse liability in humans. Lastly,
cranioplastic cement, and the guide cannulae were enclosed
the neurochemical effects of olanzapine on extracellular
within a small plastic chamber to prevent access by the
dopamine in the ventral striatum of conscious monkeys
monkeys. Stainless-steel stylets were placed in the guide
were determined with in vivo microdialysis techniques.
cannulae when not in use. Monkeys were allowed to recover
Given the recent clinical interest in potential interactions
from surgery for 2 weeks before initiating microdialysis
between selective serotonin reuptake inhibitors (SSRIs) and
experiments. All animals received Banamine (1.0 mg/kg)
atypical antipsychotics, the effects of coadministration of
every 6 for 24 h postoperatively, or longer if they exhibited
fluoxetine and olanzapine on extracellular dopamine also
were determined. Olanzapine suppressed cocaine self-administration at a dose that enhanced dopamine release,
but did not exhibit abuse liability or overt motor sideeffects. Moreover, pretreatment with fluoxetine had no
Cocaine HCl (National Institute on Drug Abuse, Rockville,
systematic effect on olanzapine-induced increases in extra-
MD) and fluoxetine HCl (Eli Lilly and Company) were
dissolved in 0.9% saline. Drug doses were determined assalts. Olanzapine (Eli Lilly and Company) was dissolved in0.01 N HCl and diluted with distilled water to appropriate
Subjects. Six female and four male adult rhesus monkeys(Macaca mulatta) weighing 7.5–13.0 kg were used as
During behavioral testing, each monkey was seated in a
subjects. Each subject was housed individually and fed
commercially available primate chair (Primate Products,
Purina monkey chow, fruits, and vegetables. Water was
Redwood City, CA, USA), and a response panel with one
continuously available. Animal care procedures strictly
lever was mounted on the front of chair. Located above the
followed the NIH ‘Guide for the Care and Use of Laboratory
lever in the center of the response panel were red and white
Olanzapine and cocaine self-administrationLL Howell et al
stimulus lights. Once the monkey was seated in the chair, a
chamber. The chair limited movement of the animals
Huber needle (Access Technologies, Skokie, IL) was
and facilitated connections between the implanted probes
inserted into the venous access port. The polyvinyl-chloride
and appropriate perfusion equipment. A Lexan plate
tubing attached to the Huber needle was connected to a
positioned perpendicular to the medial plane of the body,
motor-driven syringe (Coulbourn Instruments, Allentown,
just above shoulder height, ensured that animals could not
PA) located outside of the chamber containing the drug
contact the probe area. A microinjection pump (CMA/102)
solution. A volume of 2.0 ml/infusion was delivered over 7 s.
located outside the chamber continuously delivered artifi-
Testing during daily 1-h sessions occurred in a ventilated,
cial cerebrospinal fluid (Na2HPO4, 1.0 mM; NaCl, 150 mM;
sound-attenuating chamber. IBM compatible computers
KCl, 3 mM; CaCl, 1.3 mM; MgCl, 1.0 mM; and ascorbic acid,
controlled experimental events and recorded data.
0.15 mM) via FEP Teflon tubing to the probe for perfusion
Subjects responded for i.v. infusions of cocaine under a
second-order schedule of reinforcement, as described
During a 2-h equilibrium period, animals sat in the
previously (Wilcox et al, 2002). The training dose of
chamber, and repeated 10-min samples were obtained.
cocaine was 0.1 mg/kg/infusion. When the daily session
Subsequently, a single dose of cocaine (0.5 mg/kg), olanza-
began, the red light on the response panel was illuminated
pine (0.1 mg/kg), fluoxetine (3.0 mg/kg) or a combination of
and responding resulted in the delivery of a drug infusion
olanzapine (0.1 mg/kg) and fluoxetine (3.0 mg/kg) was
and brief 2-s illumination of the white light. Initially, the
administered i.v. to determine drug-induced increases in
fixed ratio (FR) was one (FR 1) and gradually increased to
extracellular dopamine. In drug-combination studies,
FR 20. Ultimately, a second-order schedule of reinforcement
fluoxetine was administered first, and olanzapine was
was in effect, with the first FR 20 completed after 10 min
administered 30 min later. The order of drug testing was
(fixed-interval, FI 10) resulting in a drug infusion. FR 20
randomized and counterbalanced across treatment condi-
components completed within the 10-min FI resulted in
tions, and at least 1 week separated repeated determina-
illumination of the white light for 2 s. There was a 30-s
tions. Samples were collected outside the test chamber at
limited hold for completion of the first FR 20 after the FI 10
10-min intervals to ensure that the monkeys were not
had elapsed, and a drug infusion was not delivered if the
disturbed during the experiments. Probes were tested in
limited hold expired. Drug infusions were signaled by a
vitro to determine the suitability of probe efficiency and
change in the lights from red to white for 15 s. Following
performance before and after in vivo experiments. Micro-
each drug infusion there was a 1-min timeout during which
bore HPLC and electrochemical detection quantitated
responding on the lever had no programmed consequences.
extracellular levels of dopamine according to well-estab-
A total of five infusions could be delivered during a daily
lished analytical procedures (Church et al, 1987; Skirboll
session comprising five FI 10-min (FR 20:s) components.
et al, 1990; Parsons and Justice, 1993).
The training sequence remained in effect until responding
for cocaine was stable (o20% variance in daily response
rate over five consecutive days), after which saline wassubstituted for cocaine until responding decreased to below
Response rates for individual subjects were analyzed as a
30% of responding for the training dose of cocaine. After
function of dose for each drug. Average rates of cocaine-
saline extinction, the maintenance dose (0.1 mg/kg/infu-
maintained responding following pretreatment drugs were
sion) of cocaine was reinstated and responding was allowed
compared to rates of cocaine-maintained responding
to stabilize. For pretreatment studies, a given dose of drug
following saline pretreatments using one-way ANOVA. Post
was administered i.v. 15 min presession on three consecu-
hoc comparisons were made using Dunnet’s method.
tive days, typically Tuesday, Wednesday, and Thursday.
Changes in percent basal dopamine were calculated for
Vehicle was administered on all days that subjects did not
each drug treatment (increase in extracellular dopamine in
receive a drug pretreatment, and these data contributed to
the presence of drug/basal level of dopamine) *100%. Mean
ongoing calculations of baseline stability. Pretreatment
percent basal dopamine was analyzed as a function of drug
doses were administered on two separate occasions in an
and time, and data were derived from individual subjects.
ascending order. All doses of a particular drug were studiedin combination with 0.1 mg/kg/infusion cocaine first. Subsequently, the maintenance dose of cocaine was changed
to 0.3 mg/kg/infusion, and drug pretreatments were re-
peated as described above. For drug-substitution studies,each subject was allowed to self-administer several doses of
Dose–effect curves for cocaine self-administration generally
olanzapine in a randomized order. Substitution for each
were characterized as inverted U-shaped functions in
drug dose continued for at least five consecutive sessions,
or until responding stabilized (o20% variance in daily
(0.03 mg/kg/infusion) maintained peak rates of responding
in two subjects (RMm-4 and ROu-4) (Figure 1). The highestunit dose (1.0 mg/kg/infusion) maintained rates of respond-
ing well below peak rates in all subjects. Maintenance dosesof cocaine (0.03–0.3 mg/kg/infusion) for drug pretreatment
At the time of testing, each monkey was seated in a
studies were selected to encompass doses that maintained
commercially available primate chair (Primate Products,
high, stable rates of responding above saline extinction
Inc., Miami, FL). Daily sessions lasted for approximately 4 h
levels. Pretreatment with olanzapine (0.03–0.3 mg/kg)
and were conducted within a ventilated, sound-attenuating
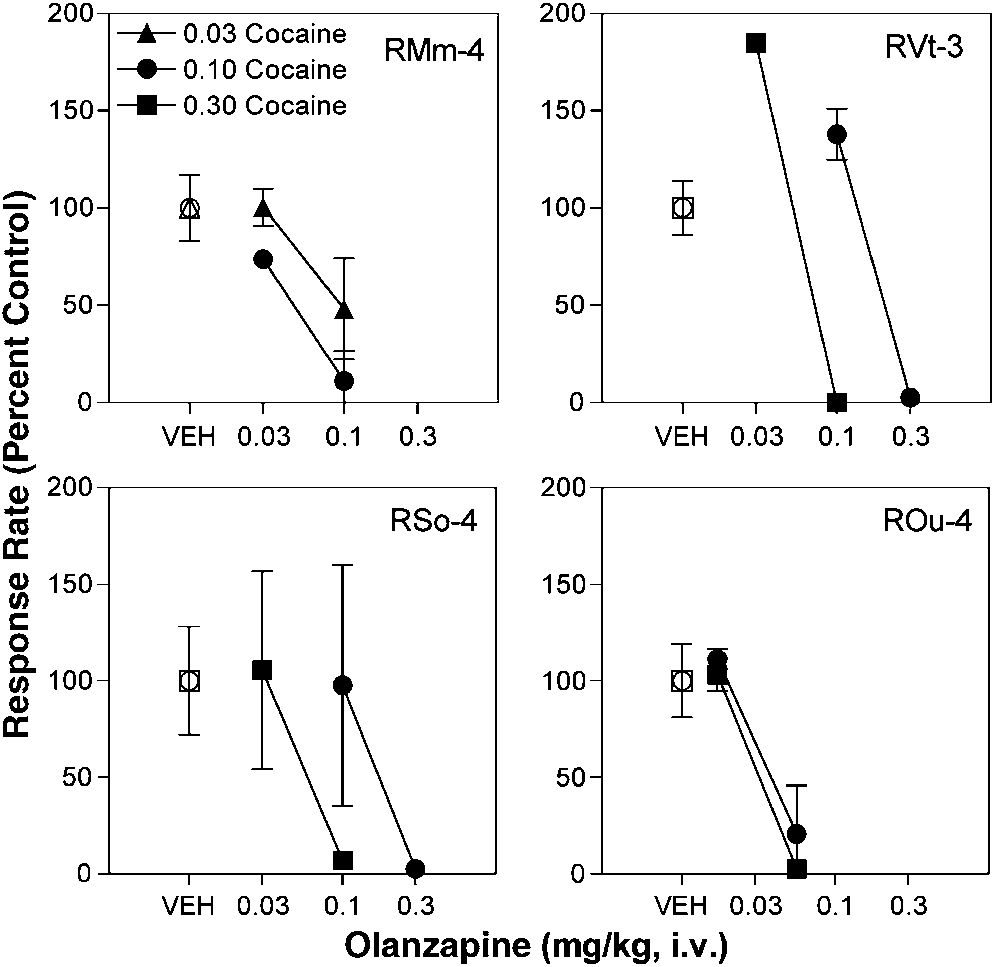
produced significant (po0.05) reductions in cocaine
Olanzapine and cocaine self-administration
Response rates (percent control) for three maintenance doses
of cocaine (0.03, 0.10, and 0.30 mg/kg/infusion) after pretreatment witholanzapine as a function of olanzapine dose under a second-order scheduleof reinforcement in rhesus monkeys. Each panel represents an individualmonkey. Data points over VEH indicate response rates when a vehiclepretreatment was given prior to the cocaine self-administration session.
The effects of olanzapine were compared directly to those
of the neuroleptic, haloperidol (Figure 3). Pretreatment witheither 0.01 mg/kg (RMk-4 and RKs-4) or 0.03 mg/kg (ROu-4) produced significant reductions in cocaine self-adminis-tration. In contrast to olanzapine pretreatments, the effectsof haloperidol were greater on the second and third days ofpretreatment compared to the first day of pretreatment. Moreover, doses of haloperidol that suppressed cocaineself-administration caused marked sedation and catalepsy. Gross observations in the animal home room after testsessions indicated pronounced effects on motor behavior. Note that lower doses of haloperidol were ineffective in
Mean (7SD) rates of responding (responses/s) maintained by
reducing cocaine self-administration and did not induce
cocaine self-administration in individual subjects. Data were derived from
observable effects on motor behavior (data not shown).
the last 5 days of each condition that met stability criterion of o20%variation. Dashed lines indicate the upper limit for responding during saline
Drug doses shown in Figure 3 represent the lowest doses
that suppressed cocaine self-administration in individualsubjects.
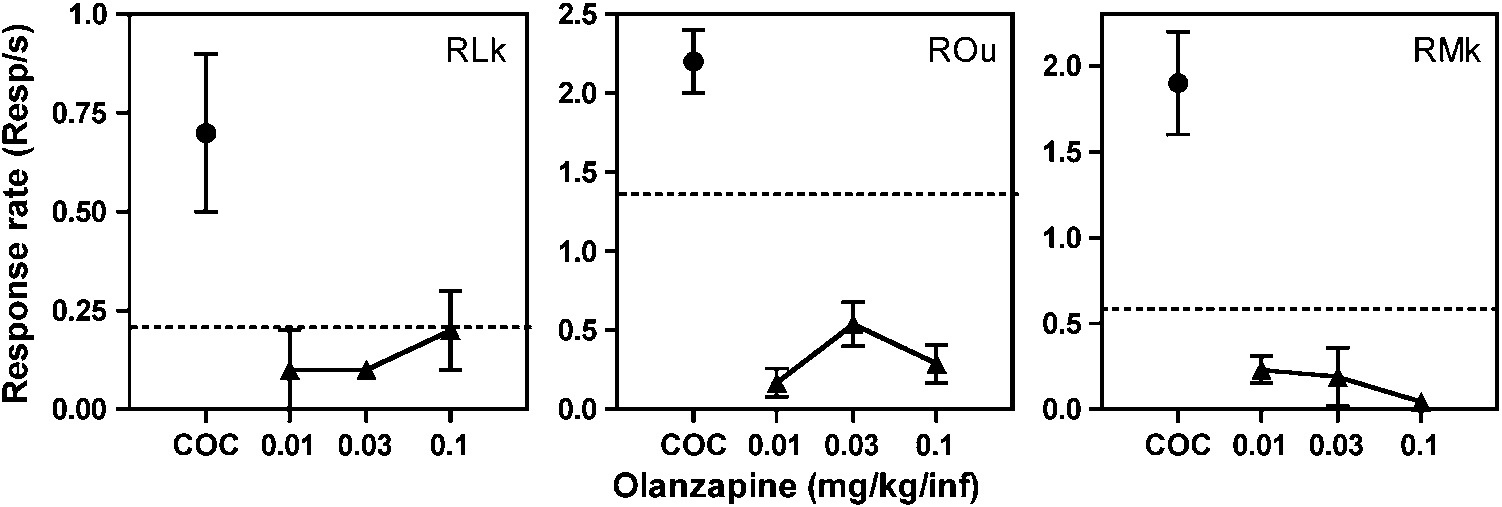
Olanzapine (0.01–0.1 mg/kg/infusion) also was substi-
self-administration in all subjects (Figure 2). There was no
tuted for cocaine in drug self-administration studies in
change in effectiveness over consecutive 3-day pretreat-
order to characterize its reinforcing effects (Figure 4). When
ments, so data were pooled across all pretreatment sessions.
different unit doses of olanzapine were substituted for
Responding was completely eliminated in three of four
cocaine for a minimum of 5 consecutive sessions, response
subjects (RVt-3, RSo-4, and ROu-4) and markedly sup-
rates were similar to or less than those maintained under
pressed in the fourth subject (RMm-4). Olanzapine was
saline substitution (extinction) conditions. The latter results
more effective in suppressing cocaine self-administration at
clearly indicated that olanzapine lacked reinforcing effects.
the higher maintenance dose of cocaine in three of foursubjects (RMm-4, RVt-3, and RSo-4) as shown by the
leftward position of the olanzapine dose–effect curves. Doses of olanzapine that produced significant reductions
Drug-induced changes in extracellular dopamine in ventral
in cocaine self-administration had no overt behavioral
striatum were characterized in awake subjects sitting quietly
effects indicative of sedation. Gross observations in the
in a primate chair. Cocaine (0.5 mg/kg) produced marked
animal home room after test sessions indicated normal
elevations in extracellular dopamine to approximately 270%
of baseline values in a group of three subjects (Figure 5).
Olanzapine and cocaine self-administrationLL Howell et al
Response rates (percent control) for two maintenance doses of cocaine (0.10 and 0.30 mg/kg/infusion) after pretreatment with haloperidol
(0.01 mg/kg, RMk-3 and RKs-4; 0.03 mg/kg, ROu-4) as a function of pretreatment day under a second-order schedule of reinforcement in rhesus monkeys. Each panel represents an individual monkey. Data points over VEH indicate response rates when a vehicle pretreatment was given prior to the cocaine self-administration session.
Response rates (responses/s) for olanzapine as a function of dose under a second-order schedule of reinforcement in rhesus monkeys. Each
panel represents an individual monkey. Data points over COC indicate response rates for the baseline cocaine dose (0.10 mg/kg/infusion). Dashed linesindicate the upper limit of responding during saline extinction. Each data point is the mean of the last five sessions in a condition.
Peak effects were observed 10–30 min postinjection and
behavior maintained by a complex, second-order schedule
returned to baseline within 60–70 min. Compared to
of i.v. drug delivery in nonhuman primates. Pretreatment
cocaine, the effects of olanzapine were less pronounced.
with olanzapine completely eliminated cocaine self-admin-
Olanzapine (0.1 mg/kg) increased extracellular dopamine to
istration in three of four subjects, and markedly suppressed
approximately 190% of baseline in the same group of three
responding to less than 10% of control rates in the fourth
subjects (Figure 5). Moreover, it had a slow onset of action
subject. Effective doses of olanzapine were within the
compared to cocaine. Peak effects were observed 60–70 min
therapeutic dose range for treatment of psychosis in
postinjection and returned to baseline within 110–120 min.
humans (Brown et al, 1999; Albers et al, 2005). In separate
Olanzapine (0.1 mg/kg) was administered in combination
experiments, substitution of olanzapine for cocaine failed to
with fluoxetine (3.0 mg/kg) to characterize their interaction
maintain self-administration behavior above saline extinc-
on neurochemistry in vivo in two subjects (Figure 6). The
tion levels in any subject. Hence, there was no indication in
dose of fluoxetine selected was based on published studies
nonhuman primates with a history of cocaine use that
reporting interactions with cocaine on behavior in nonhu-
olanzapine has abuse liability. A dose of olanzapine that
man primates (Howell and Byrd, 1995). Administration of
significantly suppressed cocaine self-administration also
fluoxetine alone had no effect on extracellular dopamine.
induced moderate increases in extracellular dopamine in
Moreover, pretreatment with fluoxetine had no systematic
the ventral striatum of conscious subjects. Lastly, the
effect on olanzapine-induced increases in extracellular
olanzapine-induced increases in dopamine were not influ-
dopamine. There was an attenuation of olanzapine-induced
enced systematically by the SSRI, fluoxetine.
increases in extracellular dopamine in one subject (RBp-3)
Doses of olanzapine that suppressed cocaine self-admin-
and an enhancement in the other subject (RJj-7).
istration had no overt behavioral effects indicative ofsedation as evidenced by gross observations in the animalhome room after test sessions. In contrast, reductions incocaine self-administration induced by the neuroleptic,
haloperidol, were accompanied by profound sedation and
This is the first study to characterize the effects of
catalepsy. Moreover, the effects of haloperidol were
olanzapine on cocaine self-administration behavior and in
enhanced over multiple pretreatment days, indicating an
vivo neurochemistry in nonhuman primates. Olanzapine
accumulation of drug effects on behavior with repeated
was effective in suppressing cocaine self-administration
dosing. It is unclear whether a more sensitive baseline of
Olanzapine and cocaine self-administration
Effects of cocaine (0.5 mg/kg) or olanzapine (0.1 mg/kg) on extracellular dopamine levels in ventral striatum of awake rhesus monkeys (n ¼ 3).
The 10-min sampling intervals indicated began 90 min after probe insertion. Data points over BL1-3 indicate three 10-min samples prior to druginjection.
Olanzapine and cocaine self-administrationLL Howell et al
zapine for cocaine in self-administration experimentsmaintained rates of responding that were actually lowerthan saline extinction conditions in two of three subjects. Inthe only other study to report the effects of an atypicalantipsychotic on cocaine self-administration in nonhumanprimates, clozapine was marginally effective in blocking thereinforcing effects of cocaine under a fixed-ratio schedule,and a high dose of clozapine decreased cocaine self-administration in an apparently nonspecific manner (Van-over et al, 1993). Hence, olanzapine may have producednonspecific disruptions in operant behavior in the presentstudy that were not evident in gross observations of home-cage behavior.
The dose of olanzapine that significantly suppressed
cocaine self-administration behavior induced a moderateincrease in extracellular dopamine measured in the ventralstriatum of conscious subjects. In the primate, the patternof limbic input to striatum includes areas of the ventralcaudate and medial ventral putamen outside that which hastraditionally been defined as the nucleus accumbens(Selemon and Goldman-Rakic, 1985; Lynd-Balta and Haber,1994; Haber and McFarland, 1999). Hence, the region ofinterest targeted in the present study may be broadlydefined as mesolimbic striatum (Bradberry et al, 2000). Compared to a behaviorally relevant dose of cocaine, whichcorresponded to the total dose administered during dailytest sessions, olanzapine-induced increases in dopaminewere less pronounced. Olanzapine also had a slower onsetand longer duration of action. The results obtained areconsistent with those reported previously in rodentsshowing olanzapine-induced increases in extracellulardopamine in nucleus accumbens (Li et al, 1998). However,it is important to note that olanzapine increases bothdopamine and norepinephrine in nucleus accumbens,striatum, and prefrontal cortex (Li et al, 1998). Moreover,olanzapine and other atypical antipsychotics induce greaterincreases in dopamine and norepinephhrine in the pre-frontal cortex compared to subcortical areas in rodents (Liet al, 1998; Westerink et al, 1998; Kuroki et al, 1999) andnonhuman primates (Youngren et al, 1999). This prefer-ential augmentation of dopamine release in prefrontalcortex may be an important aspect of their antipsychoticeffects (Li et al, 1998; Youngren et al, 1999). Accordingly,the contribution of olanzapine-induced increases in dopa-
Effects of olanzapine alone (0.10 mg/kg), fluoxetine alone
mine in ventral striatum to its effects on cocaine self-
(3.0 mg/kg), or the combination on extracellular dopamine levels in ventral
administration remains to be defined.
striatum of awake rhesus monkeys (n ¼ 2). The 10-min sampling intervals
Pretreatment with the SSRI, fluoxetine, had no systematic
indicated began 90 min after probe insertion. Data points over BL1-3
effect on olanzapine-induced increases in striatal dopamine
indicate three 10-min baseline samples prior to drug injection.
at a dose of fluoxetine that was approximately one log unithigher than the effective therapeutic dose in the treatmentof depression in humans (Wernicke et al, 1989). Similarly,
operant behavior may have detected nonspecific motor
fluoxetine failed to modulate olanzapine-induced increases
effects induced by olanzapine. Meil and Schechter (1997)
in dopamine in the striatum or nucleus accumbens of rats
reported olanzapine-induced decreases in cocaine self-
(Koch et al, 2004). It should be noted that combined
administration in rodents, but similar decreases in food-
administration of fluoxetine and olanzapine in rats
maintained behavior were observed, suggesting a nonspe-
produced sustained increases in dopamine in prefrontal
cific disruption of operant behavior. In the present study,
cortex that were significantly greater than either drug alone
olanzapine was more effective at the higher maintenance
(Zhang et al, 2000). Hence, interactions between fluoxetine
dose of cocaine in three of four subjects as shown by
and olanzapine on dopaminergic function may be regionally
the leftward position of the olanzapine dose–effect curves.
selective. Recently, there has been major interest in
The latter results are not consistent with a competitive
potential interactions between SSRIs and atypical antipsy-
pharmacological antagonism. Also, substitution with olan-
chotics, including olanzapine. Conventional antidepressant
Olanzapine and cocaine self-administration
treatment is ineffective in a significant number of patients
Brown CS, Markowitz JS, Moore TR, Parker NG (1999). Atypical
with major depression, and when concomitant psychiatric
antipsychotics: Part II: Adverse effects, drug interactions, and
symptoms are present, response outcomes are less favor-
costs. Ann Pharmacother 33: 210–217.
able. Olanzapine may provide an augmentation strategy for
Bymaster FP, Calligaro DO, Falcone JF, Marsh RD, Moore NA,
treatment-resistant and psychotic depression as evidenced
Tye NC et al (1996). Radioreceptor binding profile of the
by recent clinical studies (Thase, 2002; Shelton et al, 2001).
atypical antipsychotic olanzapine. Neuropsychopharmacology14: 87–96.
Presently, the neurochemical basis underlying the thera-
Bymaster FP, Rasmussen K, Calligaro DO, Nelson DL, DeLapp
peutic effects of fluoxetine/olanzapine combinations is
NW, Wong DT et al (1997). In vitro and in vivo biochemistry
unknown. It is reasonable to speculate that enhanced
of olanzapine: a novel, atypical antipsychotic drug. J Clin
dopamine release in prefrontal cortex may provide an
important neurochemical mechanism (Zhang et al, 2000).
Church WH, Justice Jr JB, Byrd LD (1987). Extracellular dopamine
However, the present results in conjunction with those
in rat striatum following uptake inhibition by cocaine,
reported by Koch et al (2004) suggest that augmented
nomifensine and benztropine. Eur J Pharmacol 139: 345–348.
dopamine release in mesolimbic striatum may not con-
De Wit H, Wise RA (1977). Blockade of cocaine reinforcement in
tribute to therapeutic effectiveness of fluoxetine/olanzapine
rats with the dopamine receptor blocker pimozide, but not with
the noradrenergic blockers phentolamine or phenoxybenzamine. Can J Psychol 31: 195–203.
In summary, olanzapine was effective in suppressing
Farren CK, Hameedi FA, Rosen MA, Woods S, Jatlow P, Kosten TR
cocaine self-administration behavior in nonhuman pri-
(2000). Significant interaction between clozapine and cocaine in
mates. Effective doses of olanzapine did not exhibit obvious
cocaine addicts. Drug Alcohol Depend 59: 153–163.
motor side effects or reinforcing effects indicative of abuse
Haber SN, McFarland NR (1999). The concept of the ventral
liability. A dose of olanzapine that suppressed cocaine self-
striatum in nonhuman primates. Ann NY Acad Sci 877:
administration in all subjects also induced modest increases
in mesolimbic dopamine. Although the results suggest a
Hand TH, Hu XT, Wang RY (1987). Differential effects of acute
potential therapeutic role for olanzapine in the treatment of
clozapine and haloperidol on the activity of ventral tegmental
cocaine abuse, placebo-controlled clinical studies in human
(A10) and nigrostriatal (A9) dopamine neurons. Brain Res 415:
cocaine users were not encouraging (Kampman et al, 2003).
Heikkila RE, Manzino L (1984). Behavioral properties of GBR
It remains to be determined whether adverse side effects
12909, GBR 13069 and GBR 13098: specific inhibitors of
will limit the use of olanzapine in the treatment of cocaine
dopamine uptake. Eur J Pharmacol 103: 241–248.
abuse, or whether combination therapy with SSRIs may
Hemby SE, Smith JE, Dworkin SI (1996). The effects of eticlopride
augment the effectiveness of olanzapine as a cocaine
and naltrexone on responding maintained by food, cocaine,
heroin and cocaine/heroin combinations in rats. J PharmacolExp Ther 277: 1247–1258.
Howell LL, Byrd LD (1995). Serotonergic modulation of the
behavioral effects of cocaine in the squirrel monkey. J Pharmacol
Howell LL, Wilcox KM (2001). The dopamine transporter and
This work was supported by Investigator Initiated Contracts
cocaine medication development: drug self-administration in
with Eli Lilly and Company, an Independent Scientist
nonhuman primates. J Pharmacol Exp Ther 298: 1–6.
Award (K02 DA00517) from the National Institute on Drug
Kampman KM, Pettinati H, Lynch KG, Sparkman T, O’Brien CP
Abuse, National Institutes of Health, and Grant RR00165
(2003). A pilot trial of olanzapine for the treatment of cocaine
from the Division of Research Resources, National Insti-
dependence. Drug Alcohol Depend 70: 29–37.
tutes of Health. The Yerkes National Primate Research
Kleven MS, Woolverton WL (1993). Effects of three monoamine
Center is fully accredited by the Association for the
uptake inhibitors on behavior maintained by cocaine or
Assessment and Accreditation of Laboratory Animal Care
food presentation in rhesus monkeys. Drug Alcohol Depend
International (AAALAC International). We gratefully ac-
Koch S, Perry KW, Bymaster FP (2004). Brain region and dose
knowledge the technical assistance of Dr George Keshelava,
effects of an olanzapine/fluoxetine combination on extracellular
Tango Howard, Olga Epstein, Lisa Neidert, and Peggy Plant.
monoamine concentrations in the rat. Neuropharmacology 46:232–242.
Kuhar MJ, Ritz MC, Boja JW (1991). The dopamine hypothesis
of the reinforcing properties of cocaine. Trends Neurosci 14:299–302.
Albers LJ, Ozdemir V, Marder SR, Raggi MA, Aravagiri M,
Kuroki T, Meltzer HY, Ichikawa J (1999). Effects of antipsychotic
Endrenyi L et al (2005). Low-dose fluvoxamine as an adjunct to
drugs on extracellular dopamine levels in rat medial pre-
reduce olanzapine therapeutic dose requirements: a prospective
frontal cortex and nucleus accumbens. J Pharmacol Exp Ther
dose-adjusted drug interaction strategy. J Clin Psychopharmacol
Li X-M, Perry KW, Wong DT, Bymaster FP (1998). Olanzapine
Bergman J, Madras BK, Johnson SE, Spealman RD (1989). Effects
increases in vivo dopamine and norepinephrine release in rat
of cocaine and related drugs in nonhuman primates. III. Self-
prefrontal cortex, nucleus accumbens and striatum. Psychophar-
administration by squirrel monkeys. J Pharmacol Exp Ther 251:
Lynd-Balta E, Haber SN (1994). The organization of midbrain
Bradberry CW, Barrett-Larimore RL, Jatlow P, Rubino SR (2000).
projections to the ventral striatum in the primate. Neuroscience
Impact of self-administered cocaine and cocaine cues on
extracellular dopamine in mesolimbic and sensorimotor stria-
Meil WM, Schechter MD (1997). Olanzapine attenuates the
tum in rhesus monkeys. J Neurosci 20: 3874–3883.
reinforcing effects of cocaine. Eur J Pharmacol 340: 17–26.
Olanzapine and cocaine self-administrationLL Howell et al
Moore NA, Calligaro DO, Wong DT, Bymaster F, Tye NC (1993).
effects of cocaine and dopamine transporter occupancy. Nature
The pharmacology of olanzapine and other new antipsychotic
agents. Curr Opin Invest Drugs 2: 281–293.
Wernicke JF, Bosomworth JC, Ashbrook E. (1989). Fluoxetine at
Moore NA, Rees G, Sanger G, Tye NC (1994). Effects of olanzapine
20 mg per day: the recommended and therapeutic dose in the treat-
and other antipsychotic agents on responding maintained by a
ment of depression. Int Clin Psychopharmacol 4(Suppl 1): 63–67.
conflict schedule. Behav Pharmacol 5: 196–202.
Westerink BH, de Boer P, de Vries JB, Kruse CG, Long SK (1998).
Parsons LH, Justice Jr JB (1993). Perfusate serotonin increases
Antipsychotic drugs induce similar effects on the release of
extracellular dopamine in the nucleus accumbens as measured
dopamine and noradrenaline in the medial prefrontal cortex of
by in vivo microdialysis. Brain Res 606: 195–199.
the rat brain. Eur J Pharmacol 361: 27–33.
Reith MEA, Meisler BE, Sershen H, Lajtha A (1986). Structural
Wilcox KM, Paul IA, Woolverton WL (1999). Comparison between
requirements for cocaine congeners to interact with dopamine
dopamine transporter affinity and self-administration potency
and serotonin uptake sites in mouse brain and to induce
of local anesthetics in rhesus monkeys. Eur J Pharmacol 367:
stereotyped behavior. Biochem Pharmacol 35: 1123–1129.
Ritz MC, Lamb RJ, Goldberg SR, Kuhar MJ (1987). Cocaine
Wilcox KM, Lindsey KP, Votaw JR, Goodman MM, Martarello L,
receptors on dopamine transporters are related to self-admin-
Carroll FI et al (2002). Self-administration of cocaine and the
istration of cocaine. Science 237: 1219–1223.
cocaine analog RTI-113: relationship to dopamine transporter
Selemon LD, Goldman-Rakic PS (1985). Longitudinal topography
occupancy determined by PET neuroimaging in rhesus mon-
and interdigitation of corticostriatal projections in the rhesus
Wise RA (1984). Neural mechanisms of the reinforcing action of
Shelton RC, Tollefson GD, Tohen M, Stahl S, Gannon KS,
cocaine. NIDA Res Monogr 50: 15–33.
Jacobs TG et al (2001). A novel augmentation strategy
Wise RA (1998). Drug-activation of brain reward pathways. Drug
for treating resistant major depression. Am J Psychiatry 158:
Woolverton WL (1986). Effects of a D1 and a D2 dopamine
Skirboll S, Wang J, Mefford I, Hsiao J, Bankiewicz KS (1990). In
antagonist on the self-administration of cocaine and piribedil by
vivo changes of catecholamines in hemiparkinsonian monkeys
rhesus monkeys. Pharmacol Biochem Behav 24: 531–535.
measured by microdialysis. Exp Neurol 110: 187–193.
Youngren KD, Inglis FM, Pivirotto PJ, Jedema HP, Bradberry CW,
Thase ME (2002). What role do atypical antipsychotic drugs
Goldman-Rakic PS et al (1999). Clozapine preferentially
have in treatment-resistant depression? J Clin Psychiatry 63:
increases dopamine release in the rhesus monkey prefrontal
cortex compared with the caudate nucleus. Neuropsychophar-
Vanover KE, Piercey MF, Woolverton WL (1993). Evaluation of the
reinforcing and discriminative stimulus effects of cocaine in
Zhang W, Perry KW, Wong DT, Potts BD, Bao J, Tollefson GD et al
combination with (+)-AJ76 or clozapine. J Pharmacol Exp Ther
(2000). Synergistic effects of olanzapine and other antipsychotic
agents in combination with fluoxetine on norepinephrine and
Volkow ND, Wang GJ, Fischman MW, Foltin RW, Fowler JS,
dopamine release in rat prefrontal cortex. Neuropsychopharma-
Abumrad NN et al (1997). Relationship between subjective
Safety Data Sheet: Hydrochloric Acid HCl 32% (food-industry grade) Under the guidance of the EC: 91/155/EEC, the circular of the Ministry of Industry and Trade 28/2010/TT-BCT and Vietnamese standards (TCVN 5507:2002). Identification of the product and the company Product name: Hydrochloric Acid HCl 32% (food-industry grade) Trade name: Hydrochloric Acid HCl 32% (food-industry
SCHEDA DATI DI SICUREZZA – AGRIZEB 80 1) IDENTIFICAZIONE DELLA SOSTANZA / PREPARATO E DEL FORNITORE Denominazione Commerciale: Telefono: 0471/976598 2) COMPOSIZIONE/INFORMAZIONE SUI COMPONENTI Principio MANGANESE ETHYLENBIS (DITHIOCARBAMMATE) (POLYMERIC) COMPLEX WITH ZINC SALT. 3) IDENTIFICAZIONE DEI PERICOLI Classificazione di pericolosità: Indicazioni di pericolosit�
 Neuropsychopharmacology (2005), 1–9& 2005 Nature Publishing Group
Olanzapine-Induced Suppression of CocaineSelf-Administration in Rhesus Monkeys
Leonard L Howell*,1,2, Kristin M Wilcox1, Kimberly P Lindsey1 and Heather L Kimmel1
Yerkes National Primate Research Center, Emory University, Atlanta, GA, USA; 2Department of Psychiatry and Behavioral Sciences,
The neuropharmacological profile of the atypical antipsychotic, olanzapine, is consistent with a potentially useful medication for cocaine
abuse. The present study utilized an i.v. drug self-administration paradigm in nonhuman primates to obtain definitive evidence regarding
the effectiveness of olanzapine to modulate the reinforcing effects of cocaine. The effects of olanzapine were compared directly to those
of the neuroleptic, haloperidol. Rhesus monkeys (n ¼ 7) were trained to self-administer cocaine (0.03–0.3 mg/kg/injection) under a
second-order, fixed-interval 600-s schedule with fixed ratio 20 components. Experimental sessions comprised five consecutive fixed
intervals, each followed by a 1-min timeout. In drug-interaction experiments, a single dose of olanzapine (0.03–0.3 mg/kg) or haloperidol
(0.01–0.03 mg/kg) was administered i.v. 15 min presession for at least three consecutive sessions. In drug-substitution experiments,
different doses of olanzapine (0.01–0.1 mg/kg/injection) were substituted for cocaine until responding stabilized. Olanzapine caused dose-
related decreases in cocaine self-administration at pretreatment doses that had no overt behavioral effects indicative of sedation. A dose
of 0.1 mg/kg eliminated cocaine self-administration in all subjects. In contrast, doses of haloperidol that suppressed cocaine self-
administration induced marked sedation and catalepsy. Olanzapine failed to maintain self-administration behavior above saline extinction
levels over a range of unit doses. In vivo microdialysis experiments in a second group of awake rhesus monkeys (n ¼ 3) confirmed
previous reports in rodents that olanzapine effectively increases extracellular dopamine in ventral striatum. The dose of olanzapine that
markedly suppressed cocaine self-administration behavior increased dopamine to approximately 190% of control values. Lastly,
pretreatment with fluoxetine had no systematic effect on olanzapine-induced increases in striatal dopamine. The results indicate that
olanzapine can effectively suppress cocaine self-administration behavior in nonhuman primates at doses that enhance dopamine release
but do not maintain drug self-administration.
Neuropsychopharmacology (2005), 1–9& 2005 Nature Publishing Group
Olanzapine-Induced Suppression of CocaineSelf-Administration in Rhesus Monkeys
Leonard L Howell*,1,2, Kristin M Wilcox1, Kimberly P Lindsey1 and Heather L Kimmel1
Yerkes National Primate Research Center, Emory University, Atlanta, GA, USA; 2Department of Psychiatry and Behavioral Sciences,
The neuropharmacological profile of the atypical antipsychotic, olanzapine, is consistent with a potentially useful medication for cocaine
abuse. The present study utilized an i.v. drug self-administration paradigm in nonhuman primates to obtain definitive evidence regarding
the effectiveness of olanzapine to modulate the reinforcing effects of cocaine. The effects of olanzapine were compared directly to those
of the neuroleptic, haloperidol. Rhesus monkeys (n ¼ 7) were trained to self-administer cocaine (0.03–0.3 mg/kg/injection) under a
second-order, fixed-interval 600-s schedule with fixed ratio 20 components. Experimental sessions comprised five consecutive fixed
intervals, each followed by a 1-min timeout. In drug-interaction experiments, a single dose of olanzapine (0.03–0.3 mg/kg) or haloperidol
(0.01–0.03 mg/kg) was administered i.v. 15 min presession for at least three consecutive sessions. In drug-substitution experiments,
different doses of olanzapine (0.01–0.1 mg/kg/injection) were substituted for cocaine until responding stabilized. Olanzapine caused dose-
related decreases in cocaine self-administration at pretreatment doses that had no overt behavioral effects indicative of sedation. A dose
of 0.1 mg/kg eliminated cocaine self-administration in all subjects. In contrast, doses of haloperidol that suppressed cocaine self-
administration induced marked sedation and catalepsy. Olanzapine failed to maintain self-administration behavior above saline extinction
levels over a range of unit doses. In vivo microdialysis experiments in a second group of awake rhesus monkeys (n ¼ 3) confirmed
previous reports in rodents that olanzapine effectively increases extracellular dopamine in ventral striatum. The dose of olanzapine that
markedly suppressed cocaine self-administration behavior increased dopamine to approximately 190% of control values. Lastly,
pretreatment with fluoxetine had no systematic effect on olanzapine-induced increases in striatal dopamine. The results indicate that
olanzapine can effectively suppress cocaine self-administration behavior in nonhuman primates at doses that enhance dopamine release
but do not maintain drug self-administration. Olanzapine and cocaine self-administration
and use of compounds that target dopaminergic systems
Animals’ and were approved by the Institutional Animal
represents a reasonable approach for the pharmacological
Care and Use Committee of Emory University.
Olanzapine and cocaine self-administration
and use of compounds that target dopaminergic systems
Animals’ and were approved by the Institutional Animal
represents a reasonable approach for the pharmacological
Care and Use Committee of Emory University. Olanzapine and cocaine self-administrationLL Howell et al
stimulus lights. Once the monkey was seated in the chair, a
chamber. The chair limited movement of the animals
Huber needle (Access Technologies, Skokie, IL) was
and facilitated connections between the implanted probes
inserted into the venous access port. The polyvinyl-chloride
and appropriate perfusion equipment. A Lexan plate
tubing attached to the Huber needle was connected to a
positioned perpendicular to the medial plane of the body,
motor-driven syringe (Coulbourn Instruments, Allentown,
just above shoulder height, ensured that animals could not
PA) located outside of the chamber containing the drug
contact the probe area. A microinjection pump (CMA/102)
solution. A volume of 2.0 ml/infusion was delivered over 7 s.
Olanzapine and cocaine self-administrationLL Howell et al
stimulus lights. Once the monkey was seated in the chair, a
chamber. The chair limited movement of the animals
Huber needle (Access Technologies, Skokie, IL) was
and facilitated connections between the implanted probes
inserted into the venous access port. The polyvinyl-chloride
and appropriate perfusion equipment. A Lexan plate
tubing attached to the Huber needle was connected to a
positioned perpendicular to the medial plane of the body,
motor-driven syringe (Coulbourn Instruments, Allentown,
just above shoulder height, ensured that animals could not
PA) located outside of the chamber containing the drug
contact the probe area. A microinjection pump (CMA/102)
solution. A volume of 2.0 ml/infusion was delivered over 7 s.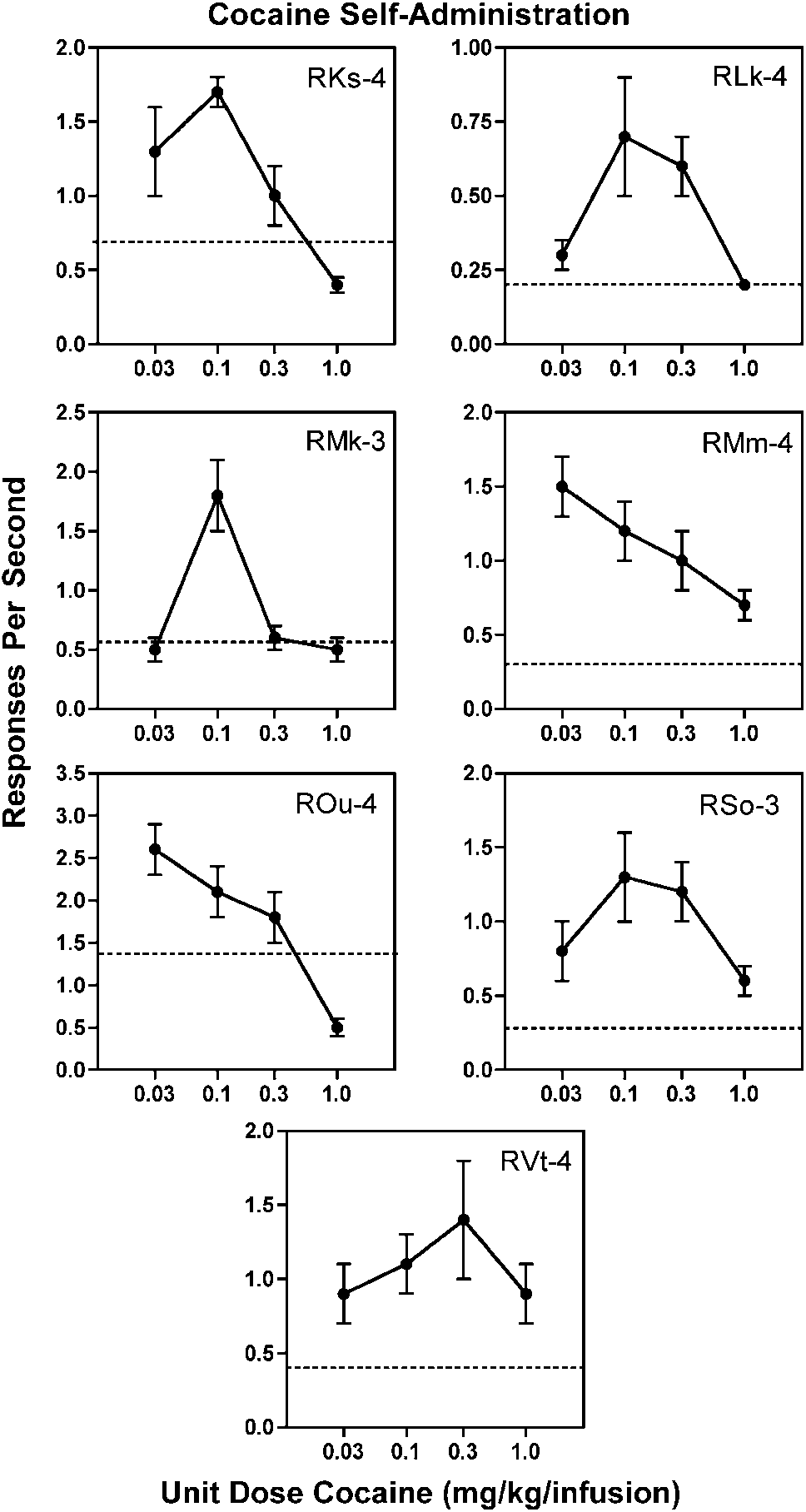
 Olanzapine and cocaine self-administration
Response rates (percent control) for three maintenance doses
of cocaine (0.03, 0.10, and 0.30 mg/kg/infusion) after pretreatment witholanzapine as a function of olanzapine dose under a second-order scheduleof reinforcement in rhesus monkeys. Each panel represents an individualmonkey. Data points over VEH indicate response rates when a vehiclepretreatment was given prior to the cocaine self-administration session.
Olanzapine and cocaine self-administration
Response rates (percent control) for three maintenance doses
of cocaine (0.03, 0.10, and 0.30 mg/kg/infusion) after pretreatment witholanzapine as a function of olanzapine dose under a second-order scheduleof reinforcement in rhesus monkeys. Each panel represents an individualmonkey. Data points over VEH indicate response rates when a vehiclepretreatment was given prior to the cocaine self-administration session.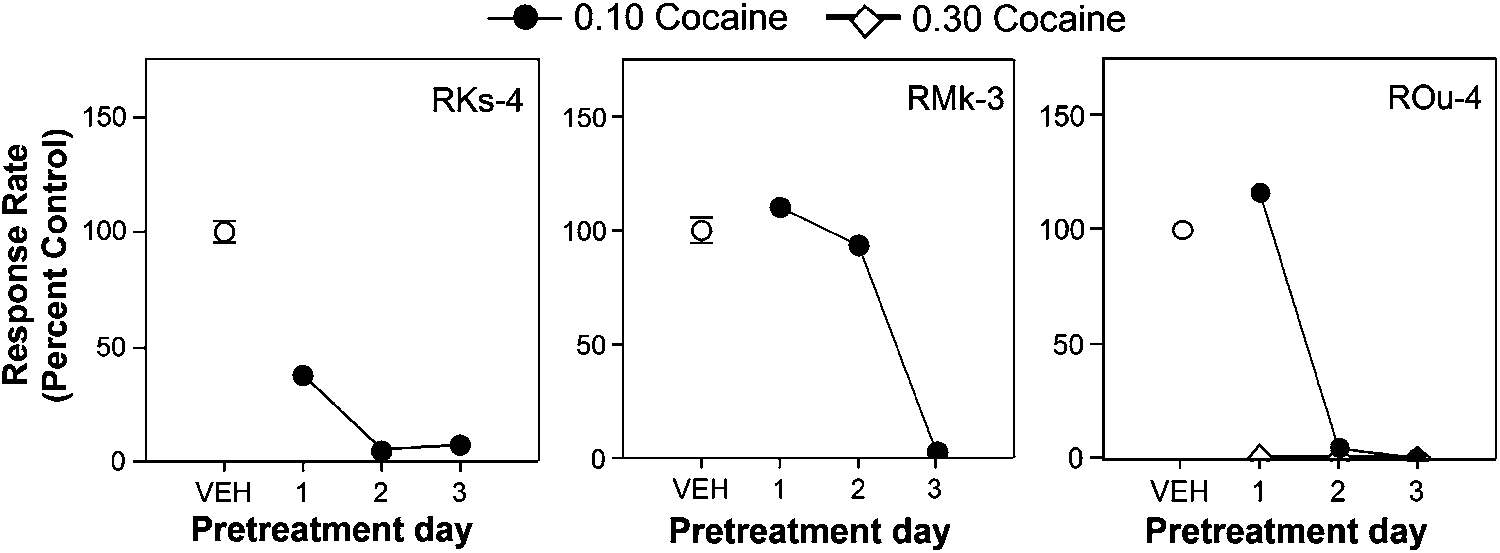
 Olanzapine and cocaine self-administrationLL Howell et al
Response rates (percent control) for two maintenance doses of cocaine (0.10 and 0.30 mg/kg/infusion) after pretreatment with haloperidol
(0.01 mg/kg, RMk-3 and RKs-4; 0.03 mg/kg, ROu-4) as a function of pretreatment day under a second-order schedule of reinforcement in rhesus monkeys.
Olanzapine and cocaine self-administrationLL Howell et al
Response rates (percent control) for two maintenance doses of cocaine (0.10 and 0.30 mg/kg/infusion) after pretreatment with haloperidol
(0.01 mg/kg, RMk-3 and RKs-4; 0.03 mg/kg, ROu-4) as a function of pretreatment day under a second-order schedule of reinforcement in rhesus monkeys.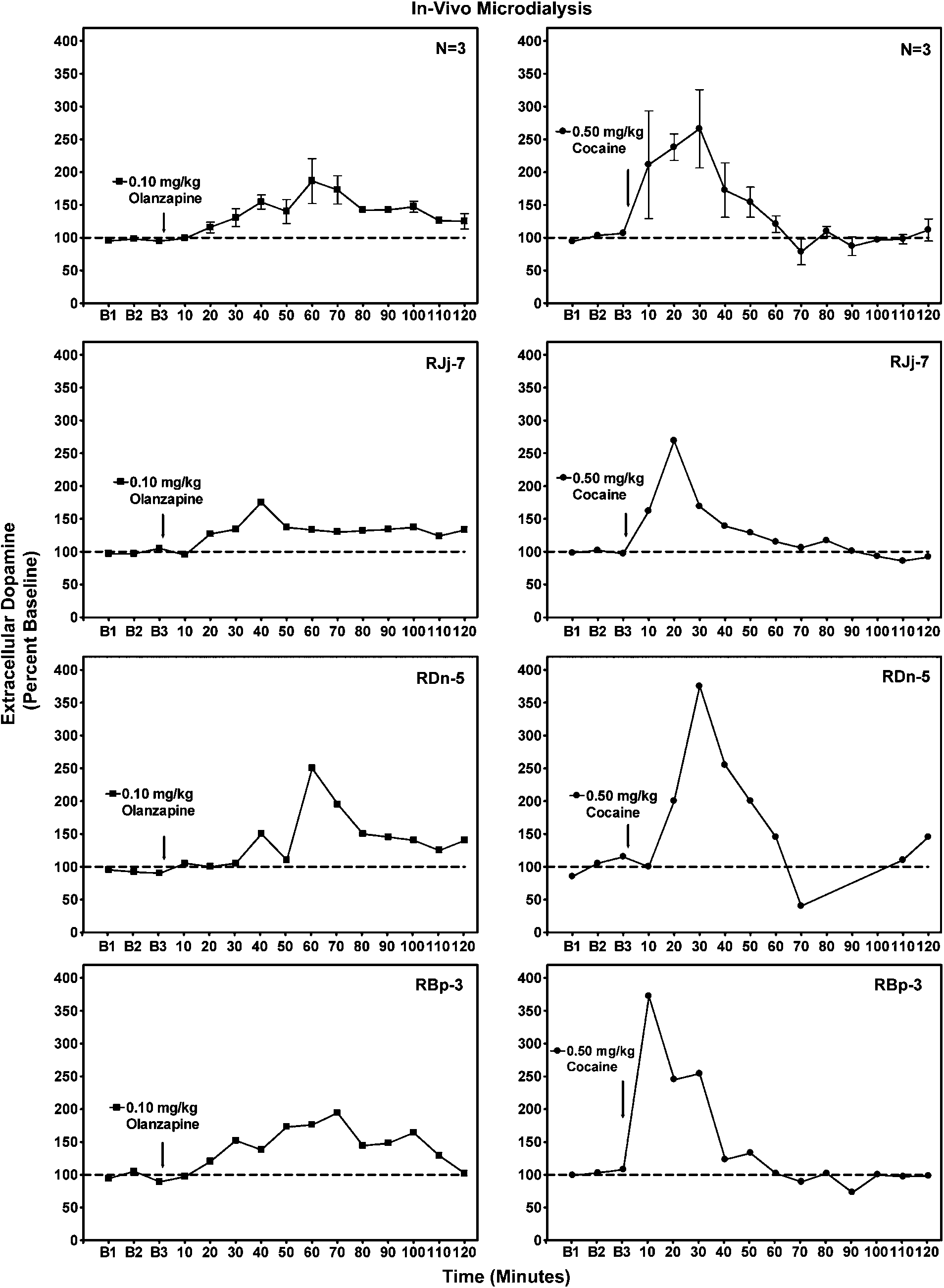
 Olanzapine and cocaine self-administration
Effects of cocaine (0.5 mg/kg) or olanzapine (0.1 mg/kg) on extracellular dopamine levels in ventral striatum of awake rhesus monkeys (n ¼ 3).
Olanzapine and cocaine self-administration
Effects of cocaine (0.5 mg/kg) or olanzapine (0.1 mg/kg) on extracellular dopamine levels in ventral striatum of awake rhesus monkeys (n ¼ 3).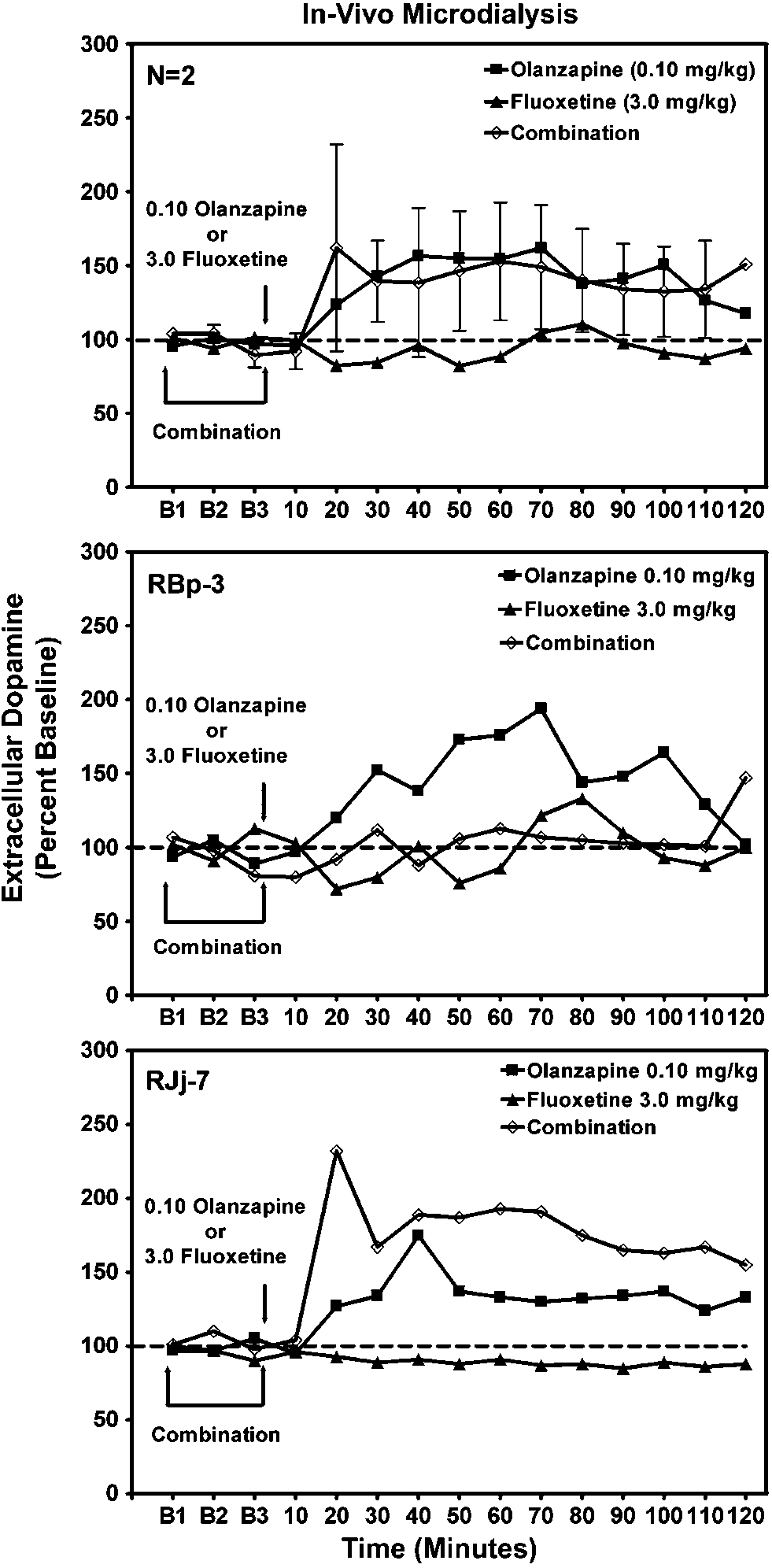
 Olanzapine and cocaine self-administrationLL Howell et al
zapine for cocaine in self-administration experimentsmaintained rates of responding that were actually lowerthan saline extinction conditions in two of three subjects. Inthe only other study to report the effects of an atypicalantipsychotic on cocaine self-administration in nonhumanprimates, clozapine was marginally effective in blocking thereinforcing effects of cocaine under a fixed-ratio schedule,and a high dose of clozapine decreased cocaine self-administration in an apparently nonspecific manner (Van-over et al, 1993). Hence, olanzapine may have producednonspecific disruptions in operant behavior in the presentstudy that were not evident in gross observations of home-cage behavior.
Olanzapine and cocaine self-administrationLL Howell et al
zapine for cocaine in self-administration experimentsmaintained rates of responding that were actually lowerthan saline extinction conditions in two of three subjects. Inthe only other study to report the effects of an atypicalantipsychotic on cocaine self-administration in nonhumanprimates, clozapine was marginally effective in blocking thereinforcing effects of cocaine under a fixed-ratio schedule,and a high dose of clozapine decreased cocaine self-administration in an apparently nonspecific manner (Van-over et al, 1993). Hence, olanzapine may have producednonspecific disruptions in operant behavior in the presentstudy that were not evident in gross observations of home-cage behavior. Olanzapine and cocaine self-administration
treatment is ineffective in a significant number of patients
Brown CS, Markowitz JS, Moore TR, Parker NG (1999). Atypical
with major depression, and when concomitant psychiatric
antipsychotics: Part II: Adverse effects, drug interactions, and
symptoms are present, response outcomes are less favor-
costs. Ann Pharmacother 33: 210–217.
Olanzapine and cocaine self-administration
treatment is ineffective in a significant number of patients
Brown CS, Markowitz JS, Moore TR, Parker NG (1999). Atypical
with major depression, and when concomitant psychiatric
antipsychotics: Part II: Adverse effects, drug interactions, and
symptoms are present, response outcomes are less favor-
costs. Ann Pharmacother 33: 210–217. Olanzapine and cocaine self-administrationLL Howell et al
Moore NA, Calligaro DO, Wong DT, Bymaster F, Tye NC (1993).
Olanzapine and cocaine self-administrationLL Howell et al
Moore NA, Calligaro DO, Wong DT, Bymaster F, Tye NC (1993).