Simulated Docking of Zanamivir with the 2009 Pandemic Strain Influenza A/H1N1 Neuraminidase Active Site Abstract Influenza neuraminidases are glycoproteins that facilitate the transmission of the influenza virus from cell to cell. Zanamivir is a widely used neuraminidase inhibitor. Here I provide a computational docking analysis of zanamivir with the active site of the neuraminidase of the 2009 Influenza A/H1N1 strain. The computed inhibitor/receptor binding energy suggests that zanamivir would be only marginally effective against that strain. Keywords: Influenza, H1N1, neuraminidase, zanamivir
1.0 Introduction
glycoproteins that facilitate the transmission
the group-1 N1, N4 and N8 neuraminidases
of the influenza virus from cell to cell.
([1]) reveal that the active sites of these
dimensional structure from that of group-2
enzymes. The differences lie in a loop of
amino acids known as the "150-loop", which
conformation that opens a cavity not present
pyran-2-carboxylic acid; [10]) is a widely
loop contains an amino acid designated Asp
151; the side chain of this amino acid has a
carboxylic acid that, in group-1 enzymes,
points away from the active site as a result
of the 'open' conformation of the 150-loop.
related sero-subtypes, and these subtypes
The side chain of another active-site amino
correspond at least roughly to differences in
conformation in group-1 enzymes compared
neuraminidases. The subtypes fall into two
with the group-2 neuraminidases (8]).
groups ([3]): group-1 contains the subtypes
acid side chains form critical interactions
with neuraminidase inhibitors. For neuraminidase subtypes with the “open
conformation” 150-loop, the side chains
is a structural description of most of the
inhibitors tightly ([8]). The active site
([10]) using AutoDock Tools v 4.2 (ADT,
docking of zanamivir to the receptor. More
outside the active site. This means that
against another with the same active-site
extracted. (3TI3 identifies the active site of
with some group 2-like features in its active
torsions in the ligand and active site were
2.0 Method
to be flexible wherever that assumption is
assess the binding energy of the active site
physically possible, was auto-docked to the
of crystallized A/California/04/2009(H1N1))
active site, assumed to be rigid, using the
Lamarckian genetic algorithm implemented
otherwise noted, all processing described in
configuration from the analysis was saved,
Inspiron 545 with an Intel Core2 Quad CPU
and the distances between the receptor and
ligand in 3TI3, and those computed here,
RAM, running under the Windows Vista
docking are shown in Figure 1. Most values are, or are a consequence of, ADT defaults.
________________________________________________________________________ autodock_parameter_version 4.2 # used by autodock to validate parameter set outlev 1 # diagnostic output level intelec # calculate internal electrostatics seed pid time # seeds for random generator ligand_types C HD OA N # atoms types in ligand fld 3TI3_active.maps.fld # grid_data_file map 3TI3_active.C.map # atom-specific affinity map map 3TI3_active.HD.map # atom-specific affinity map map 3TI3_active.OA.map # atom-specific affinity map map 3TI3_active.N.map # atom-specific affinity map elecmap 3TI3_active.e.map # electrostatics map desolvmap 3TI3_active.d.map # desolvation map move zanamivir.pdbqt # small molecule about -29.5772 12.7517 -20.6465 # small molecule center tran0 random # initial coordinates/A or random axisangle0 random # initial orientation dihe0 random # initial dihedrals (relative) or random tstep 2.0 # translation step/A qstep 50.0 # quaternion step/deg dstep 50.0 # torsion step/deg torsdof 9 # torsional degrees of freedom rmstol 2.0 # cluster_tolerance/A extnrg 1000.0 # external grid energy e0max 0.0 10000 # max initial energy; max number of retries ga_pop_size 150 # number of individuals in population ga_num_evals 2500000 # maximum number of energy evaluations ga_num_generations 27000 # maximum number of generations ga_elitism 1 # number of top individuals to survive to next generation ga_mutation_rate 0.02 # rate of gene mutation ga_crossover_rate 0.8 # rate of crossover ga_window_size 10 # ga_cauchy_alpha 0.0 # Alpha parameter of Cauchy distribution ga_cauchy_beta 1.0 # Beta parameter Cauchy distribution set_ga # set the above parameters for GA or LGA sw_max_its 300 # iterations of Solis & Wets local search sw_max_succ 4 # consecutive successes before changing rho sw_max_fail 4 # consecutive failures before changing rho sw_rho 1.0 # size of local search space to sample sw_lb_rho 0.01 # lower bound on rho ls_search_freq 0.06 # probability of performing local search on individual set_psw1 # set the above pseudo-Solis & Wets parameters unbound_model bound # state of unbound ligand ga_run 10 # do this many hybrid GA-LS runs analysis # perform a ranked cluster analysis
Figure 1. ADT parameters for the docking in this study
______________________________________________________________________________ 3.0 Results
processor (with occasional bursts to 40% of
peak), and required a constant 2.9 GB of memory.
which assumes familiarity with the general
neuraminidase "landscape", took about 20
minutes in ADT; the docking proper, about
energy of binding under these conditions is
~ -8.7 kcal/mol; the estimated inhibition
monitor suggested that the calculation was
more or less uniformly distributed across the four processors at ~25% of peak per ______________________________________________________________________________ MODEL 1 USER Run = 1 USER Cluster Rank = 1 USER Number of conformations in this cluster = 10 USER USER RMSD from reference structure = 56.144 A USER USER Estimated Free Energy of Binding = -8.72 kcal/mol [=(1)+(2)+(3)-(4)] USER Estimated Inhibition Constant, Ki = 408.13 nM (nanomolar) [Temperature = 298.15 K] USER USER (1) Final Intermolecular Energy = -11.40 kcal/mol USER vdW + Hbond + desolv Energy = -8.30 kcal/mol USER Electrostatic Energy = -3.10 kcal/mol USER (2) Final Total Internal Energy = -2.75 kcal/mol USER (3) Torsional Free Energy = +2.68 kcal/mol USER (4) Unbound System's Energy [=(2)] = -2.75 kcal/mol USER USER USER USER DPF = 3TI3_zanamivir.dpf USER NEWDPF move zanamivir.pdbqt USER NEWDPF about -29.577200 12.751700 -20.646500 USER NEWDPF tran0 29.961176 14.781299 -20.419074 USER NEWDPF axisangle0 -0.004045 -0.391949 0.919978 3.081993 USER NEWDPF quaternion0 -0.000109 -0.010540 0.024740 0.999638 USER NEWDPF dihe0 4.89 175.54 139.90 180.00 67.18 1.07 -179.74 0.58 -36.96 USER USER x y z vdW Elec q RMS ATOM 1 C2 ZMR A1001 29.610 13.398 -22.778 -0.14 +0.09 +0.144 56.144 ATOM 2 C3 ZMR A1001 30.901 13.720 -22.564 -0.34 +0.01 +0.045 56.144 ATOM 3 C4 ZMR A1001 31.277 14.664 -21.442 -0.27 -0.00 +0.150 56.144 ATOM 4 C5 ZMR A1001 30.226 14.586 -20.317 -0.17 +0.04 +0.143 56.144 ATOM 5 C6 ZMR A1001 28.817 14.747 -20.891 -0.14 +0.08 +0.185 56.144 ATOM 6 O6 ZMR A1001 28.541 13.810 -21.924 -0.14 -0.22 -0.335 56.144 ATOM 7 NE ZMR A1001 32.576 14.369 -20.810 -0.22 +0.04 -0.217 56.144 ATOM 8 HE ZMR A1001 32.843 13.389 -20.711 -0.26 -0.16 +0.178 56.144 ATOM 9 CZ ZMR A1001 33.401 15.265 -20.371 +0.01 +0.06 +0.665 56.144 ATOM 10 NH1 ZMR A1001 33.240 16.579 -20.493 -0.24 +0.05 -0.235 56.144 ATOM 11 NH2 ZMR A1001 34.493 14.843 -19.724 -0.31 -0.14 -0.235 56.144 ATOM 12 2HH1 ZMR A1001 32.407 16.900 -20.987 +0.08 -0.07 +0.174 56.144 ATOM 13 1HH1 ZMR A1001 33.890 17.285 -20.148 -0.38 -0.08 +0.174 56.144 ATOM 14 2HH2 ZMR A1001 34.617 13.835 -19.630 -0.39 +0.16 +0.174 56.144 ATOM 15 1HH2 ZMR A1001 35.144 15.549 -19.378 -0.44 +0.11 +0.174 56.144 ATOM 16 N5 ZMR A1001 30.437 15.627 -19.309 -0.02 -0.20 -0.352 56.144 ATOM 17 H5 ZMR A1001 30.130 16.576 -19.525 +0.10 +0.07 +0.163 56.144 ATOM 18 C10 ZMR A1001 31.013 15.406 -18.112 -0.24 +0.22 +0.214 56.144 ATOM 19 C11 ZMR A1001 31.268 16.657 -17.329 -0.34 +0.13 +0.117 56.144 ATOM 20 O10 ZMR A1001 31.344 14.278 -17.729 -0.74 -0.41 -0.274 56.144 ATOM 21 C1 ZMR A1001 29.129 12.658 -23.951 -0.19 +0.35 +0.233 56.144 ATOM 22 O1A ZMR A1001 30.010 12.129 -24.683 -1.05 -1.46 -0.642 56.144 ATOM 23 O1B ZMR A1001 27.908 12.571 -24.177 -1.03 -1.48 -0.642 56.144 ATOM 24 C7 ZMR A1001 27.690 14.594 -19.863 -0.09 +0.13 +0.180 56.144 ATOM 25 C8 ZMR A1001 26.561 15.617 -20.084 -0.25 +0.09 +0.173 56.144 ATOM 26 O8 ZMR A1001 25.343 14.887 -20.303 -0.20 -0.19 -0.391 56.144 ATOM 27 H8 ZMR A1001 24.662 15.515 -20.514 -0.40 -0.11 +0.210 56.144 ATOM 28 C9 ZMR A1001 26.902 16.556 -21.266 -0.21 +0.02 +0.198 56.144 ATOM 29 O9 ZMR A1001 25.780 16.637 -22.140 -0.01 -0.06 -0.398 56.144 ATOM 30 H9 ZMR A1001 25.104 16.044 -21.835 -0.35 -0.03 +0.209 56.144 ATOM 31 O7 ZMR A1001 27.148 13.287 -19.968 +0.01 -0.32 -0.390 56.144 ATOM 32 H7 ZMR A1001 27.094 13.052 -20.887 +0.08 +0.19 +0.210 56.144 TER ENDMDL
Figure 2. ADT's zanamivir energy and position predictions. ______________________________________________________________________________
Figure 3 is a rendering of the active-site/inhibitor configuration computed in this study. Figure 3. Rendering of zanamivir computationally docked with the active site of PDB 3TI3. The molecular surface of the receptor is shown in white; the inhibitor, in stick form in grey. Only the interior, inhibitor-containing region of the molecular surface of the active site can be compared to in situ data: the surface distal to the interior is a computational artifact, generated by the assumption that active site is detached from the rest of the receptor. ______________________________________________________________________________ The distances between ligand and receptor
however, be more effective than oseltamivir
distances in the present computation were
assumes that the receptor is rigid. This
assumption is appropriate for the binding
4.0 Discussion
However, the calculation does not reflect
what receptor "flexing" could contribute to
The method described in Section 2.0 and the
the interaction of the ligand with native
Sections 2.0 and 3.0 assumes receptor is in a
computed in this study (~408 nanoMolar at
crystallized form. In situ, at physiologically
~298 K) is comparable inhibition constant of
normal temperatures (~310 K), the receptor
clinically effective ([10], [11], [13], [14],
[15]) against several H1N1 genotypes. This
therefore, may not be identical to their
genetic algorithm used in this work could be
[5] Butler D. Avian flu special: The flu
models could be applied to this problem, and
pandemic: were we ready? Nature 435 (26
its own active site. The work described in
this paper was performed on Chain A only.
highly similar to the Chain A active site.
Future work will assess the ligand/receptor
Summary: Interim Recommendations for the Use of Influenza Antiviral Medications in the Setting of Oseltamivir Resistance among Circulating Influenza A (H1N1) Viruses, 5.0 Acknowledgements 2008-09 Influenza Season. 19 December 2008.
This work benefited from discussions with
[8] Luo M. Structural biology: antiviral
drugs fit for a purpose. Nature 443 (7
6.0 References.
[1] Russell RJ et al. The structure of H5N1
opportunities for drug design. Nature 443 (6
the accounts: global mortality of the 1918-1920 "Spanish " influenza pandemic.
Bulletin of the History of Medicine 76
X, Zhu X, Dwek RA, Stevens J, Wilson IA.
[3] World Health Organization. A revision
Journal of Virology 82 (2008), 10493-
of the system of nomenclature for influenza
viruses: a WHO memorandum. Bulletin of the World Health Organization 58 (1980),
efficacies of RWJ-270201, oseltamivir, and
avian influenza viruses. Antimicrobial
M, Liu Y, Gao F, Liu J, Feng E, He J, Wang
Agents and Chemotherapy 45 (2001), 2723-
J, Liu H, Jiang H, and Gao GF. Structural
and functional analysis of laninamivir and
its octanoate prodrug reveals group specific
AutoDock 4 with AutoDock Tools: A
neuraminidase active site. Proceedings of the 2011 International Conference on Genetic and Evolutionary Methods. CSREA
Relationship between the inhibition constant
(Ki) and the concentration of inhibitor which
causes 50 per cent inhibition (I50) of an
Pharmacology 22 (December 1973), 3099–
site. Proceedings of the 2011 International Conference on Genetic and Evolutionary Methods. CSREA Press. pp. 136-142.
oseltamivir with the 1918 pandemic strain
REVIEW ARTICLE Smoking Cessation Therapy and the Return of Aviators to Flying Duty Alon Grossman, Dan-Avi Landau, Erez Barenboim,and Liav Goldstein GROSSMAN A, LANDAU D-A, BARENBOIM E, GOLDSTEIN L. Smoking who have experienced a myocardial infarction and con- cessation therapy and the return of aviators to flying duty. Aviat tinue to smoke compared with those who quit. Further-
Medical data is for informational purposes only. You should always consult your family physician, or one of our referral physicians prior to treatment. tients from angina, claudication, and dizziness. However with longterm and extensive use the benefits of chelates are lost with thenonspecific removal and reduction of the essential trace metals. Chelation therapy can be a double edge sword li
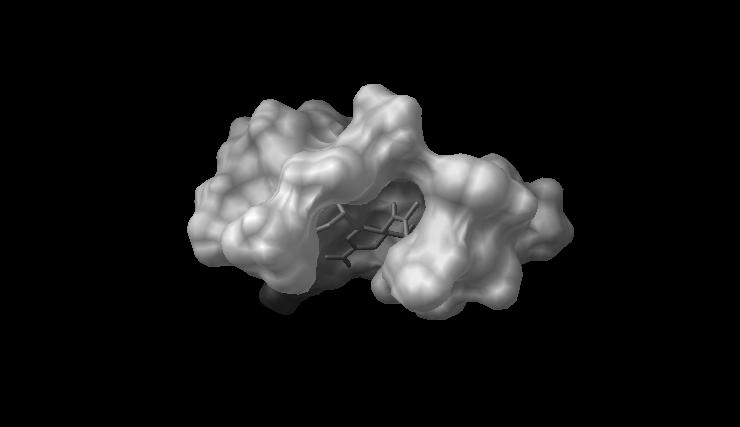 Figure 3. Rendering of zanamivir computationally docked with the active site of PDB 3TI3.
Figure 3. Rendering of zanamivir computationally docked with the active site of PDB 3TI3.