This Provisional PDF corresponds to the article as it appeared upon acceptance. Fully formatted
PDF and full text (HTML) versions will be made available soon. Inverse relation between FASN expression in human adipose tissue and the insulin resistance level Nutrition & Metabolism 2010, 7:3 Article type Submission date Acceptance date Publication date Article URL
This peer-reviewed article was published immediately upon acceptance. It can be downloaded,
printed and distributed freely for any purposes (see copyright notice below).
Articles in Nutrition & Metabolism are listed in PubMed and archived at PubMed Central.
For information about publishing your research in Nutrition & Metabolism or any BioMed Central
For information about other BioMed Central publications go to
2010 Mayas et al. , licensee BioMed Central Ltd.
This is an open access article distributed under the terms of the Creative Commons Attribution License
which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited. Inverse relation between FASN expression in human adipose tissue and the insulin resistance level
María D Mayas1,2,§, Francisco J Ortega 2,3, Manuel Macías-González2,4, Rosa Bernal
1,2, Ricardo Gómez-Huelgas4, José M Fernández-Real 2,3, Francisco J Tinahones1,2
1Servicio de Endocrinología y Nutrición, Hospital Clínico Universitario Virgen de
2CIBEROBN (CB06/03/010), Instituto de Salud Carlos III, España
3Servicio de Diabetes, Endocrinología y Nutrición, Instituto de Investigación
4Laboratorio de Investigación, Fundación IMABIS, Málaga, España
5Servicio de Medicina Interna, Hospital Universitario Carlos Haya de Málaga, España
RGH: [email protected]Abstract Background
Adipose tissue is a key regulator of energy balance playing an active role in lipid
storage and may be a dynamic buffer to control fatty acid flux. Just like PPARγ, fatty
acid synthesis enzymes such as FASN have been implicated in almost all aspects of
human metabolic alterations such as obesity, insulin resistance or dyslipemia. The aim
of this work is to investigate how FASN and PPARγ expression in human adipose
tissue is related to carbohydrate metabolism dysfunction and obesity.
The study included eighty-seven patients which were classified according to their
BMI and to their glycaemia levels in order to study FASN and PPARγ gene
expression levels, anthropometric and biochemical variables.
The main result of this work is the close relation between FASN expression level and
the factors that lead to hyperglycemic state (increased values of glucose levels,
HOMA-IR, HbA1c, BMI and triglycerides). The correlation of the enzyme with these
parameters is inversely proportional. On the other hand, PPARγ is not related to
Conclusions
We can demonstrate that FASN expression is a good candidate to study the
pathophysiology of type II diabetes and obesity in humans.
Background Adipose tissue is recognized as a key regulator of energy balance, playing an active role
in lipid storage with multiple distinct deposits (subcutaneous, intra-abdominal and
intrathoracic) [1]. Indeed, adipocytes of visceral abdominal fat origin are more
endocrinologically active than the subcutaneous variety [2]. In addition, adipose tissue
can buffer, synthesize and secrete a wide range of endocrinal products into circulating
blood that is influential on the systemic metabolism and may be directly involved in the
pathogenesis of associated complications such as obesity, diabetes, vascular damage and
atherosclerosis [1, 3]. Thus, adipose tissue may serve as a dynamic buffer to control fatty
acid (FA) flux in response to changing energy demands: in the fasting state, adipose
tissue releases FAs, whereas in the fed state, adipocytes change to “absorb” FAs from the
circulation, mainly from circulating triglycerides (TG) [4, 5]. This function is known to
be altered in obese subjects with metabolic syndrome features (insulin resistance,
obesity, dyslipemia, inflammation, atherosclerosis and hypertension) [6, 7].
The nuclear receptor peroxisome proliferator-activated receptor gamma (PPARγ) is a
ligand-activated transcription factor, member of the nuclear hormone receptor
superfamily, which functions as a heterodimer with a retinoid X receptor (RXR) [8]. The
actions of PPARγ are mediated by two protein isoforms which are derived from the same
gene by alternative promoter usage and splicing: the widely expressed PPARγ1 and the
adipose tissue-restricted PPARγ2 [9]. The activation of PPARγ leads to adipocyte
differentiation and fatty-acid storage, whereas it represses genes that induce lipolysis and
the release of free fatty acids (FFAs) in adipocytes [10]. Authors have shown that the
loss-of-function mutation of PPARγ results in severe insulin resistance and causes
elevated TG and decreased high density lipoprotein-cholesterol levels in humans while
increased PPARγ activity enhances insulin sensitivity and improves dyslipidemia in
PPARγ transcriptionally regulates many genes involved in metabolism [12], even those
involved in the synthesis of FAs. There are two sources of FA, exogenously-derived
(dietary) and endogenously-synthesized FA, both are essential constituents of biological
membrane lipids and important substrates for energy metabolism. The biosynthesis of
the latter is catalysed by Fatty Acid Synthase (FASN) and Acetyl-CoA Carboxylase
(ACC), key enzymes of lipogenesis that may play a crucial role in the weight variability
of abdominal adipose tissue [13]. Specifically, FASN (EC 2.3.1.85) is a multifunctional
enzymatic complex, important in the regulation of body weight and the development of
obesity [13-15] and necessary for de novo synthesis of long-chain saturated FAs from
acetyl coenzyme A (CoA), malonyl-CoA and NADPH. The expression of this enzyme is
highly dependent on nutritional conditions in lipogenic tissues. FASN-catalysed
endogenous FA biosynthesis in liver and adipose tissue is stimulated by a high
carbohydrate diet, whereas it is suppressed by the presence of small amounts of FA in
There are several studies that connect FASN activity/expression with metabolic
alterations in humans such as obesity, dyslipemia, insulin resistance and altered
adipocytokine serum profile [17]. Although there are authors that have shown how
FASN gene expression is significantly higher in obese vs lean individuals [17-19],
there are studies that found the way in which FASN mRNA expression was decreased
in the subcutaneous adipose tissue of obese vs lean individuals [20]. Divergent
findings may be explained by differences in metabolic parameters and the size of the
study population. We contribute to study the role of FASN with a general population
with a wide range of body mass index (BMI) and metabolic parameters, in order to
clarify the association between FASN activity/expression, the grade of insulin
resistance and obesity-related insulin resistance.
Experimental subjects
The study included 87 healthy persons (35 men and 52 women) who underwent
laparoscopic surgery procedures (hiatus hernia repair or cholecystectomies). Patients
were classified into three groups according to BMI: normal (BMI < 25), overweight (25
≤ BMI < 30) and obese (BMI ≥ 30). Patients were also classified into normoglycemic
(no diabetes antecedents and glucose levels in a fast state ≤110 mg/dl) and
hyperglycemic (diabetics or people with basal glycaemia values in a fast state >110
mg/dl) groups. This study was approved by the Hospital’s Ethical Committee and all
participants signed their consent after being fully informed of its goal and characteristics.
Study design
Before surgery and after an overnight fast, the patient’s height and weight was measured
to calculate the BMI and the waist and circumference to calculate the waist to hip ratio
(W-H). In addition, systolic blood pressure (SBP) and diastolic blood pressure (DBP)
were noted. During surgical intervention, biopsies of visceral adipose tissue were
immediately frozen in liquid nitrogen and stored at -80ºC for gene expression analysis.
Blood samples were collected; serum and plasma were separated in aliquots within 30
min of extraction, and immediately frozen at –80ºC.
Biochemical variables were: glucose, cholesterol, TG, high density lipoprotein-
cholesterol (HDL-c) and low density lipoprotein-cholesterol (LDL-c), glycated
haemoglobin (HbA1c), C-reactive protein (CRP) and all were measured in a Dimension
Autoanalyzer (Dade Behring, Deerfield, IL) in duplicate. Serum insulin concentration
was analyzed by an immunoradiometric assay (IRMA) (BioSource International,
Camarillo, CA). Leptin and adiponectin were analysed by enzyme immunoassay
(ELISA) kits (Mediagnost, Reutlingen, Germany and DRG Diagnostics GmbH,
Germany, respectively). The homeostasis model assessment of insulin resistance
(HOMA-IR) was calculated as follows: fasting glucose (mg/dl) * fasting insulin (uU/ml)
RNA extraction and real time quantitative PCR: Adipose tissue RNA isolation was
performed by homogenization with an ULTRATURRAX T25 basic (IKA Werke GmbH,
Staufen, Germany) using Trizol reagent (Invitrogen, Barcelona, Spain). Samples were
purified using RNAEasy Mini kit (QIAGEN, Barcelona, Spain) and treated with DNase
(RNase-free DNase Set, Qiagen). For first strand cDNA synthesis, constant amounts of
1µg of total RNA were reverse transcribed using random hexamers as primers and
Transcriptor Reverse Transcriptase (Roche, Mannheim, Germany). Gene expression was
assessed by real time PCR using an ABI Prism 7000 Sequence Detection System
(Applied Biosystems, Darmstadt, Germany), using TaqMan® technology suitable for
relative genetic FASN expression quantification. The reaction was performed, following
the manufacturers protocol, in a final volume of 25µl. The cycle program consisted of an
initial denaturing of 10min at 95ºC, followed by 40 15sec denaturizing phase cycles at
95ºC and a 1min annealing and extension phase at 60ºC. Commercially available and
pre-validated TaqMan® primer/probe sets were used as follows: PPIA (4333763, RefSeq. NM_002046.3, Cyclophilin A (PPIA), used as endogenous control for the target
gene in each reaction) and FASN (Hs00188012_m1, RefSeq. NM_004104.4, Fatty Acid Synthase). A threshold cycle (Ct value) was obtained for each amplification curve and a
∆Ct value was first calculated by subtracting the Ct value for human PPIA cDNA from
the Ct value for each sample and transcript. Fold changes compared with the endogenous
control were then determined by calculating 2-∆Ct, so FASN expression results are
expressed as the expression ratio relative to PPIA gene expression according to the
manufacturer’s guidelines. The transcript levels of nuclear receptors PPARγ1 and
PPARγ2 were quantified by real-time reverse transcription RT-PCR, using LightCycler®
technology (Roche Diagnostic, Rotkreuz, Switzerland) with SYBR Green detection. The
primers for the PCR reaction (Sigma Proligo) were: a common reverse primer for
PPARγ1 and for PPARγ2, CTTCCATTACCGAGAGATCC. The forward primer for
PPARγ1 was AAAGAAGGCGACAACTAAACC and GCGATTCCTTCACTGATAC
for PPARγ2. A standard curve was created with serial dilutions of a PCR fragment from
human adipose tissue total RNA (Clontech Laboratories, Inc., Mountain View, CA). For
quantification purposes, PPARγ mRNA levels were always reported to the levels of β-
actin, constitutively expressed gene. Primers for β-actin were
AACTGGAACGGTGAAGGTGAC as forward and TGTGGACTTGGGAGAGGACTG
as reverse. All samples were quantified in duplicate and positive and negative controls
Statistical analysis
Data are expressed as mean ± standard deviation (SD). The differences in the study
variables of normal, overweight and obese individuals were compared with an ANOVA
or Student test for independent samples. Pearson’s correlation coefficients were
calculated to estimate the linear correlations between variables and the confidence
interval was of 95%. Multiple regression analysis was used to study which variables
were associated with FASN expression levels. Values were considered to be statistically
significant when P≤0.05. The statistical analyses and graphics were performed using the
program SPSS (Version 11.5 for Windows; SPSS, Chicago; IL).
Results The anthropometric and biochemical variables of the studied subjects and FASN and
PPARγ gene expression of the three groups (normal, overweight and obese) are
summarized in Table 1. BMI is directly related to SBP values (P<0.01), W-H ratio
(P<0.05), glucose (P<0.01), HbA1c (P<0.01), HOMA-IR (P<0.01), leptin (P<0.01) and
TG (P<0.01) levels and inversely related to FASN expression (P<0.01) and adiponectin
Comparisons between normoglycemic and hyperglycemic subjects (Table 2) have shown
that the last group had significantly higher baseline TG readings (P<0.05), BMI
(P<0.05), glucose (P<0.01), HbA1c (P<0.01) and HOMA-IR (P<0.01), and lower levels
of FASN expression (P<0.01). No significant changes were detected in the other
Differences according to sex (data not shown) for clinical and laboratory data have
shown that leptin and adiponectin levels were significantly higher in females (P<0.01),
the same as CRP (P<0.05) and HDL-c (P<0.01). No differences were found in the rest of
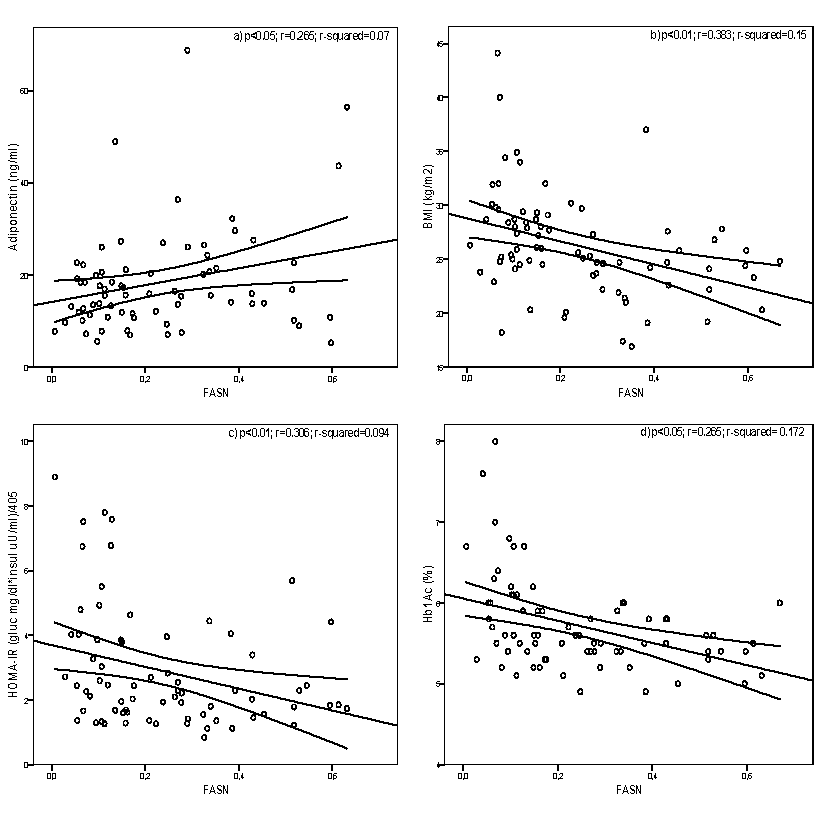
The correlation between FASN expression and the different parameters that are
associated with diabetes and obesity have shown the following results: there is a positive
correlation of FASN expression with levels of adiponectin (P<0.05; r=0.265; Figure 1a)
and HDL-c (P<0.05; r=0.276). BMI (P<0.01; r=0.383; Figure 1b), W-H (P<0.05;
r=0.274), glucose (P<0.01; r=0.373), HOMA-IR (P<0.01; r=0.306; Figure 1c), HbA1c
(P<0.01; r=0.415; Figure 1d) and TG (P<0.01; r=0.339) correlates inversely with FASN
Multiple regression analysis (Table 3) found that FASN expression levels (as dependent
variable) were related to values of HbA1c (P<0.01) and BMI (P<0.01) with a value of
the model of R2=0.385 and R2=0.271 respectively. Variables that did not enter in the
model were TG, HDL-c, HOMA-IR, adiponectin and PPARγ1 and PPARγ2 (data not
Discussion We investigated how FASN gene expression in human adipose tissue is related to
carbohydrate metabolism dysfunctions and obesity. FASN gene expression was studied
in adipose tissue using quantitative RT-PCR in samples of visceral adipose tissue from
87 volunteers who varied in terms of BMI, sex and metabolic parameters. We used
correlation analysis to dissect whether and to what extent FASN mRNA expression is
explained by the variability in anthropometric and metabolic parameters and we found an
inverse correlation of FASN with Glucose, HOMA-IR, HbA1c, TG, BMI and W-H,
while there was a positively correlation with adiponectin and HDL.
Feeding on simple carbohydrates substantially increases the activity of FASN, the central
enzyme for de novo synthesis of long-chain saturated FAs [22]. FASN expression and
activity are increased by insulin in cultured human adipocytes, suggesting that insulin
sensitivity plays a role in their regulation and is essential in the uptake of glucose and
conversion to TG. Insulin stimulates the transcription of lipogenic genes in rat
hepatocytes and adipocytes, and this action has been confirmed in human adipocytes
[23]. The results of the present study also demonstrate that adipose FASN gene
expression is higher in normoglycemic individuals compared to those with
hyperglycaemia, together with lower values of BMI, TG and obviously glucose, HOMA-
IR, and HbA1c levels in normoglycemics. The relation between FASN and glycaemia is
corroborated by multiple regression analysis where we have demonstrated the close
relation of FASN expression with HbA1c. Due to the fact that HbA1c is image of
medium values of glycaemia in the last three months, we took this value as
representative of glycaemia state. This relation is of more importance when we take into
account that what is being analyzed is a population with a wide range of BMI and
metabolic parameters. Moreover, FASN is a variable that plays a role in body weight
regulation and the development of obesity [13-15]. In this and previous studies, our
laboratory has found that FASN relates inversely with obesity and this suggests that it
could play a role in obesity-associated diabetes.
Our study design also allowed us to investigate the relationship between FASN mRNA
expression and serum concentrations of adipocytokines (leptin and adiponectin). We
found a correlation between FASN and serum concentrations of adiponectin. These
adipocytokines are also BMI dependent in obesity while leptin increases, adiponectin
decreases. According to sex we can also see that both are present in higher
concentrations in women than in men. Leptin could directly suppress FASN mRNA
expression in adipose tissue, since experimentally increased plasma leptin concentrations
in rats resulted in a decrease of FASN mRNA levels in fat [24]. There are data
supporting a suppressive action of leptin on FAS transcription [25]. Adiponectin is an
exclusively adipocyte-derived hormone [26] with a key role in glucose and lipid
metabolism in skeletal muscle and the liver, acting as an insulin sensitizer [27]. It is the
only adipocytokine known to be down-regulated in obesity [28] and insulin resistance by
decreasing TG content in muscle and liver [29]. Hypoadiponectinemia has been more
closely related to the degree of insulin resistance and hyperinsulinemia than the degree of
PPARγ has been implicated in almost all aspects of the cluster of human diseases
designated as metabolic syndrome [6, 7]. Because of this, it is a good candidate to study,
crucial for whole-body insulin sensitivity [30] and adipogenesis [8]. The actions of
PPARγ are mediated by two protein isoforms, the widely expressed PPARγ1 and the
adipose tissue-restricted PPARγ2 [9]. PPARγ also transcriptionally regulates many genes
involved in metabolism [12]. But we have found no significant changes in PPARγ1 and
PPARγ2 expression levels related to carbohydrate metabolism or FASN expression
levels. Our results do not support the relation of PPARγ with FASN and insulin
sensitivity. On the other hand, PPARγ activation is also associated with potentially
beneficial effects on the expression and secretion of adipocytokines [30] which protect
nonadipose tissue against lipid overload. Increased TNFα, leptin, and resistin levels and
decreased adiponectin expression in adipose tissues are associated with the development
of insulin resistance and vice versa [28, 30].
Conclusions Taken together, it has been demonstrated that FASN is a candidate gene for the
pathophysiology of human obesity and type II diabetes and we corroborate this with the
correlation of adipose FASN mRNA expression with several parameters related to
Competing interests The authors declare that they have no competing interests. Authors' contributions
MDM drafted the manuscript, designed the study, participated in the genetic studies
and in the analysis of biochemical variables, and performed the statistical analysis.
FJO and MMG carried out the genetic studies. RB analyzed biochemical variables.
RGH obtained the anthropometrical characteristics and the written consent of patients.
JMFR participated in the conception and the coordination of the study. FJT carried
out the conception, design and the coordination of the study, and helped with the
statistical analysis. All authors read and approved the final manuscript.
Acknowledgements This work was supported by Ministerio de Educación y Ciencia (SAF 2006/12894),
CIBEROBN (CB06/03/010), Instituto de Salud Carlos III (PI07953 and CP04/0039) and
Consejería de Innovación, Ciencia y Empresa (CTS04369).
References
1. Klaus S: Adipose tissue as a regulator of energy balance.Curr Drug Targets 2004, 5: 241-250
2. Wajchenberg BL: Subcutaneous and visceral adipose tissue: their relation to the metabolic syndrome.Endocr Rev 2000, 21:697-738
3. Grundy SM: Obesity, metabolic syndrome, and cardiovascular disease.J Clin Endocrinol Metab 2004, 89:2595-2600
4. Tsuchida A, Yamauchi T, Kadowaki T: Nuclear Receptors as Targets for Drug Development: Molecular Mechanisms for Regulation of Obesity and Insulin Resistance by Peroxisome Proliferator-Activated Receptor, CREB-Binding Protein, and Adiponectin J Pharmacol Sci 2005, 97:164- .
5. Frayn KN: Adipose tissue as a buffer for dayly lipid flux.Diabetologia
2002, 45:1201-1210
6. Scott CL: Diagnosis, prevention, and intervention for the metabolic syndrome.Am J Cardiol 2003, 92:35i-42i
7. Ginsberg HN: Treatment for patients with the metabolic syndrome.Am J Cardiol 2003, 91:29E-39E
8. Rosen ED, Spiegelman BM: PPARγ: a nuclear regulator of metabolism, differentiation, and cell growth.J Biol Chem 2001, 276:37731-37734
9. Vidal-Puig AJ, Considine RV, Jimenez-Linan M, Werman A, Pories WJ, Caro
JF, Flier JS: Peroxisome proliferator-activated receptor gene expression in human tissues. Effects of obesity, weight loss, and regulation by insulin and glucocorticoids. J Clin Invest 1997, 99:2416-2422
10. Cock TA, Houten SM, Auwerx J: Peroxisome proliferator-activated receptor-γ: too much of a good thing causes harm.EMBO reports 2004, 5
11. Barroso I, Gurnell M, Crowley VE, Agostini M, Schwabe JW, Soos MA,
Maslen GL, Williams TD, Lewis H, Schafer AJ, Chatterjee VK, O’Rahilly S:
Dominant negative mutations in human PPAR_ associated with severe insulin resistance, diabetes mellitus and hypertension.Nature 1999, 402:880-883
12. Fajas L, Debril MB, Auwerx J: Peroxisome proliferator-activated receptor- gamma: From adipogenesis to carcinogenesis.J Mol Endocrinol 2001,
13. Mobbs CV, Makimura H: Block the FAS, lose the fat.Nat Med 2002, 8:335-
14. Loftus TM, Jaworsky DE, Frehywot GL, Townsend CA, Ronnett GV, Lane
MD, Kuhajda FP: Reduced food intake and body weight in mice treated with fatty acid synthase inhibitors.Science 2000, 288:2379-2381
15. Diraison F, Dusserre E, Vidal H, Sothier M, Beylot M: Increased hepatic lipogenesis but decreased expression of lipogenic gene in adipose tissue in human obesity.Am J Physiol Endocrinol Metab 2002, 282:46-51
16. Wakil S: Fatty acid synthase, a proficient multifunctional enzyme. Biochemistry 1989, 28:4523-4530
17. Berndt J, Kovacs P, Ruschke K, Klöting N, Fasshauer M, Schön MR, Körner
A, Stumvoll M, Blüher M: Fatty acid synthase gene expression in human adipose tissue: association with obesity and type 2 diabetes.Diabetologia
2007, 50:1472-1480
18. Blüher M, Michael MD, Peroni OD, Ueki K, Carter N, Kahn BB, Kanh CR:
Adipose tissue selective insulin receptor knockout protects against obesity and obesity-related glucose intolerance.Dev Cell 2002, 3:25-38
19. Blüher M, Patti ME, Gesta S, Kahn BB, Kahn CR: Intrinsic heterogeneity in adipose tissue of fat-specific insulin receptor knock-out mice is associated with differences in patterns of gene expression. J Biol Chem 2004, 279:31891-31901
20. Turner SM, Roy S, Sul HS, Neese RA, Murphy EJ, Samandi W, Roohk DJ,
Hellerstein MK: Dissociation between adipose tissue fluxes and lipogenic gene expression in ob/ob mice.Am J Physiol Endocrinol Metab 2007, 292:E1101-E1109
21. García-Fuentes E, García-Almeida JM, García-Arnés J, Rivas-Marín J,
Gallego-Perales JL, González-Jiménez B, Cardona I, García-Serrano S,
Garriga MJ, Gonzalo M, de Adana MS, Soriguer F: The cannabinoid CB1 receptor antagonist SR141716A (Rimonabant) enhances the metabolic benefits of long-term treatment with oleoylethanolamide in Zucker rats. Neuropharmacology 2008, 54:226-34
22. Hillgartner FB, Salati LM, Goodridge AG: Physiological and molecular mechanisms involved in nutritional regulation of fatty acid synthesis. Physiol Rev 1995, 75:47-76
23. Wang Y, Jones Voy B, Urs S Kim S, Soltani-Bejnood M, Quigley N, Heo YR,
Standridge M, Andersen B, Dhar M, Joshi R, Wortman P, Taylor JW, Chun J,
Leuze M, Claycombe K, Saxton AM, Moustaid-Moussa N: The human fatty acid synthase gene and de novo lipogenesis are coordinately regulated in human adipose tissue.J Nutr 2004, 134:1032-1038
24. Nogalska A, Sucajtys-Szulc E, Swierczynski J: Leptin decreases lipogenic enzyme gene expression through modification of SREBP-1c gene expression in white adipose tissue of aging rats. Metabolism 2005, 54:1041-
25. Bai Y, Zhang S, Kim K, Lee J, Kim R: Obese gene expression alters the ability of 30A5 prreadipocytes to respond to lipogenic hormones.J Biol Chem 1996, 271:13939-13942
26. Maeda N, Takahashi M, Funahashi T, Kihara S, Nishizawa H, Kishida K,
Nagaretani H, Matsuda M, Kouro R, Ouchi N, Kuriyama H, Hotta K,
Nakamura T, Shimomura I, Matsuzawa Y: PPARgamma ligands increase expression and plasma concentrations of adiponectin, an adipose-derived protein. Diabetes 2001, 50:2094-2099
27. Chandran M, Phillips SA, Ciaraldi T, Henry RR: Adiponectin: more than just another fat cell hormone? Diabetes Care 2003, 26:2442-2450
28. Weyer C, Funahashi T, Tanaka S, Hotta K, Matsuzawa Y, Pratley RE,
Tataranni PA: Hypoadiponectinemia in obesity and type 2 diabetes: Close association with insulin resistance and hyperinsulinemia.J Clin Endocrinol Metab 2001, 86:1930-1935
29. Arita Y, Kihara S, Ouchi N, Takahashi M, Maeda K, Miyagawa J, Hotta K,
Shimomura I, Nakamura T, Miyaoka K, Kuriyama H, Nishida M, Yamashita
S, Okubo K, Matsubara K, Muraguchi M, Ohmoto Y, Funahashi T,
Matsuzawa Y: Paradoxical decrease of an adipose-specific protein, adiponectin, in obesity.Biochem Biophys Res Commun 1999, 257:79-83
30. Di Gregorio GB, Yao-Borengasser A, Rasouli N, Varma V, Lu T, Miles LM,
Ranganathan G, Peterson CA, McGehee RE, Kern PA: Expression of CD68 and macrophage chemoattractant protein-1 genes in human adipose and muscle tissues: association with cytokine expression, insulin resistance, and reduction by pioglitazone.Diabetes 2005, 54:2305-2313 Table 1 - Anthropometrical and biochemical characteristics of study subjects: normal, overweight and obese individuals
Values are presented as means ± SD. BMI, body mass index (Kg/m2); SBP, systolic
blood pressure (mm Hg); DBP, diastolic blood pressure (mm Hg); W-H ratio, waist to
hip ratio; Insulin (UI/ml); Glycaemia (mg/dl); HbA1c, glycated haemoglobin (%);
HOMA-IR, homeostasis model assessment ((gluc mg/dl*insul U/ml)/405); Cholesterol
(mg/dl); TG, triglycerides (mg/dl); LDL-c, low density lipoprotein-cholesterol (mg/dl);
HDL-c, high density lipoprotein-cholesterol (mg/dl); CRP, c-reactive protein (mg/l);
Adiponectin (ng/ml); Leptin (ng/ml); FASN, fatty acid synthase; PPARγ, peroxisome
Table 2 - Anthropometrical and biochemical characteristics of study subjects: with and without high glycaemia Patients High Glycaemia P
Values are presented as means ± SD. BMI, body mass index (Kg/m2); SBP, systolic
blood pressure (mm Hg); DBP, diastolic blood pressure (mm Hg); W-H ratio, waist to
hip ratio; Insulin (UI/ml); Glycaemia (mg/dl); HbA1c, glycated haemoglobin (%);
HOMA-IR, homeostasis model assessment ((gluc mg/dl*insul U/ml)/405);
Cholesterol (mg/dl); TG, triglycerides (mg/dl); LDL-c, low density lipoprotein-
cholesterol (mg/dl); HDL-c, high density lipoprotein-cholesterol (mg/dl); CRP, c-
reactive protein (mg/l); Adiponectin (ng/ml); Leptin (ng/ml); FASN, fatty acid
synthase; PPARγ, peroxisome proliferator-activated receptor. Relationship between
variables in control and hyperglycemic individuals was assessed by Student’s t test.
Table 3 - Multiple regression analysis
BMI, body mass index (Kg/m2); W-H, waist to hip ratio; Glucose (mg/dl); HbA1c, glycated
haemoglobin (%); HOMA-IR, homeostasis model assessment ((gluc mg/dl*insul U/ml)/405);
TG, triglycerides (mg/dl); LDL-c, low density lipoprotein-cholesterol (mg/dl); HDL-c, high
density lipoprotein-cholesterol (mg/dl); Adiponectin (ng/ml); FASN, fatty acid synthase; PPARγ,
peroxisome proliferator-activated receptor.
Dependent variable: FASN Excluded variables: PPARγ1, γ2, Adiponectin, HOMA-IR, Glucose, TG, HDL-c, W-H
Figure legend Figure 1 - Linear relationship between FASN expression and adiponectin (a), BMI (b), HOMA-IR (c) and HbA1c (d) Linear relationship was determined by Pearson’s correlation coefficient test. 95%
rendelési javasolt fogy.ár Akkordeon Standard basszus Zongorabillenyt ű s HOHNER BRAVO II 48 fekete, tokkal, Nr. 50-es hordszíjjalHOHNER BRAVO II 48 vörös, tokkal, Nr. 50-es hordszíjjalHOHNER BRAVO II 48 kék, tokkal, Nr. 50-es hordszíjjalHOHNER BRAVO II 60 piros, tokkal, Nr. 50-es hordszíjjalHOHNER BRAVO III 72 fekete, tokkal, Nr. 51-es hordszíjjalHOHNER BRAVO III 7
La Serva Padrona, de Giovanni Baptista Pergolesi (1710/1736) I – O C o m p o s i t o r Este compositor, de origem napolitana, destacou-se, tal como muitos da mesma região, pelas suas criações operísticas, que, em Nápoles, encontravam um gênero específico, o da "Ópera Buffa" (que significa ópera cômica), embora também tivesse composto obras instrumentais (ainda
 This Provisional PDF corresponds to the article as it appeared upon acceptance. Fully formatted
PDF and full text (HTML) versions will be made available soon.
This Provisional PDF corresponds to the article as it appeared upon acceptance. Fully formatted
PDF and full text (HTML) versions will be made available soon.